#生物學小講座 | 如何理解轉基因?請從這篇文章開始_風聞
Raffaele- 2018-10-10 13:45
作者:觀察者網風聞社區 Raffaele/飛揚
轉基因無疑是近年來最能引發網絡熱(hu)烈(xiang)討(si)論(bi)的科技話題,沒有之一。和政治歷史等等話題有所不同的是,正確理解轉基因需要的前置知識對於吃瓜羣眾而言,實在太多了,而僅僅介紹幾個關鍵知識點,又非常容易讓人們對轉基因產生一知半解的猜測和主觀臆斷,往往造成各種誤解誤會,讓生物學從業人員感到難以解釋清楚,無法描述明白。因此,兩位筆者決定從現代生物學最基礎,最入門的知識講起,一點一點為大家真正理解轉基因提供最充實的條件。畢竟,真正的理解才是闢謠的最佳手段。

不經歷煙熏火燎,哪裏來的火眼金睛
PART I 中心法則和基因
一、中心法則
中心法則顧名思義,是現代生物學最重要最核心的部分,無論什麼樣的生物學研究,都離不開對它的應用。那麼,什麼是中心法則呢?簡而言之,**中心法則就是遺傳信息在三種生物大分子之間週轉的形式。**哪三種生物大分子?脱氧核糖核酸DNA, 核糖核酸RNA, 蛋白質protein。這三種大分子都能承載一定的遺傳信息。按照這些週轉形式在自然界中的常見程度,我們可以得到以下的一張表格【1】

雖然中心法則自1958年被DNA的發現者克里克提出以來【2】,已經經過了極大的補充和完善,但在大多數情況下,我們需要了解的中心法則,僅僅是左側的三種遺傳信息轉換形式,即:
從DNA到DNA(稱之為DNA的自我複製)
從DNA到RNA(稱之為RNA的轉錄)
從RNA到蛋白質(稱之為蛋白質的翻譯)
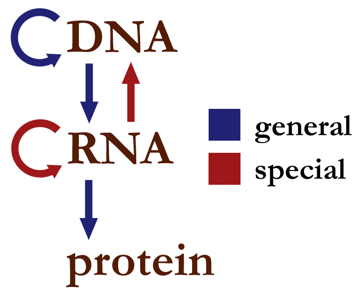
此示意圖不包括從DNA直接合成蛋白質的途徑
用一句話總結中心法則就是DNA自我複製,並轉錄成RNA,再翻譯成蛋白質,而蛋白質則協助前三個過程的進行。但當我們討論遺傳信息轉換的時候,千萬記得從DNA到RNA再到蛋白質,僅僅是真正的中心法則的一部分。
記住,只有明白了中心法則,能夠在實際問題中應用中心法則,才能説你可以理解現代生物學,尤其是和基因相關的分子生物學的內容。事實上,由於中心法則是如此的重要,筆者決定在以後的每篇相關文章中,都重複介紹它,直到吃瓜羣眾們真正把它吃透為止。
二、基因
明白了中心法則的意義之後,我們不免要問,那鋪天蓋地宣傳的基因是什麼,它和中心法則有什麼關係?在分子生物學中,基因通常被定義為一段DNA或RNA分子,是遺傳的基本物質和功能單位。為什麼基因不能是蛋白質?因為從中心法則中我們看到,蛋白質攜帶的遺傳信息基本不能轉換成DNA或者RNA,而後兩者卻沒有這個限制。

雙鏈DNA
既然基因是一段DNA或者RNA,那麼它一定也承載了一部分遺傳信息。實際上,基因(gene)這個詞,本身就是從古希臘語γόνος, gonos衍生而來,而後者在希臘語中的意義,就是生殖和後代【3】。既然基因是一個實際存在的分子,那麼它就能參與生物化學反應,這些反應,就有可能改變這個承載遺傳信息的物質基礎,也就是基因本身。這就是對基因的操作。那麼,對基因我們能進行什麼樣的操作呢?下面我們就為小白/老爺們娓娓道來。
PART II 各類基因操作方式
最近有個新聞《切開不發黑這種轉基因蘋果美國已種超100萬棵》【4】,文章中提到,美國有一種“轉基因蘋果”所打的果汁不會褐變,又提到,這種“轉基因蘋果”使用了“基因沉默”技術。可能很多讀者沒搞明白這些術語有何區別,在這裏我們就對這些術語進行一個分析。
一、轉基因transgene
我們通常所説“轉基因技術”,是指將外源基因導入目標物種並使其能夠穩定遺傳的技術【5,6】。所謂的“外源基因”,通常是目標物種(比如大豆)所不具有的基因,例如來自細菌的EPSP合成酶基因。該基因變化過程通常指那些在實驗室裏通過重組DNA技術人工插入其他物種基因以創造出新特性的手段。這些轉基因動植物與普通動植物有什麼差異呢?
1. 基因序列不同,也就是中心法則中,DNA層次上的不同。轉基因植物插入了一個它原本沒有的基因,引起基因序列的改變。
2. 表達的蛋白質不同。通過中心法則,我們知道DNA會經由RNA的轉錄,最終表達出有特定功能的蛋白質。轉入了新基因,通常就會有新的蛋白在細胞中表達。
3. 表達的其他物質不同。一部分基因並不是以表達蛋白的方式起作用,它們在轉錄成某些特殊的RNA,比如微小RNA(MicroRNA,miRNA)、長非編碼RNA(long non-coding RNA,lncRNA)、環狀RNA(circRNA)之後,就能直接參與到細胞的生命活動中去。如果轉基因動植物所插入的基因是編碼這些RNA而不是蛋白質,那麼轉入這個基因也會相應的引起這些物質發生改變。

能額外合成胡蘿蔔素的黃金大米
二、雜交hybridization
在我們談論育種的時候,雜交是指不同基因型的個體間進行的交配。通過雜交而讓雙親基因重新組合(即同源重組),讓同一種物種,甚至親緣關係相當近,且不具有生殖隔離的兩個物種進行不同個體間的繁殖,從而選出擁有優良性狀的後代。雜交通常不會引入外源基因,僅僅是親本個體間基因的交換。
網絡上很多言論認為雜交就是廣義的轉基因,甚至有更激進的觀點認為雜交就是轉基因。這種説法顯然是錯得相當離譜的。雖然在分子層面的本質上,雜交和轉基因都無非是DNA分子序列的改變,但無論從技術流程還是實際的操作結果來看,轉基因和雜交都是兩種完全不同的技術。
雜交和轉基因的區別:
1、轉基因依賴眾多分子生物學實驗手段,而雜交不需要。
2、轉基因是對外源基因的複製和轉移,而雜交是物種內部基因的重新組合。
3、轉基因能較容易的掌握新性狀的變化,而雜交不能預知同源重組的精確結果。【注1】
沒有哪一種技術是萬能的。儘管相對於雜交,轉基因技術擁有不少優勢,但就現階段而言,轉基因技術還受到相當多的限制,比如一些複雜性狀的獲得和集中,單靠轉基因技術無法實現,仍然需要依賴傳統的雜交技術。【7】【注 2】。
三、基因編輯gene editing
基因編輯技術指能夠讓人類對目標基因進行更加精確的“編輯”,實現對特定DNA片段的修改、敲除、加入等。目前較成熟的基因編輯技術包括但不限於鋅指ZFN (Zinc-Finger Nucleases)、TALEN (Transcription Activator-LikeEffector Nucleases) ,以及近年來在學術界和媒體走紅的CRISPR/Cas9技術【8】。
這個技術主要作用是什麼呢?
1、基因敲除:如果想使某個基因的功能喪失,可以通過基因編輯,在這個基因內部產生雙鏈DNA斷裂,細胞在對這類DNA損傷進行修復的過程中往往會產生DNA的隨機插入或刪除,破壞原有基因的功能,從而實現基因敲除。
2、特異突變引入:如果想把某個特異的突變引入到基因組上,需要通過同源重組來實現,這時候要提供一個含有特異突變同源模版。正常情況下同源重組效率非常低,而在這個位點產生雙鏈DNA斷裂會極大的提高重組效率,從而實現特異突變的引入。這裏涉及的原理比較複雜,我們將會專門撰文另行敍述。
3、定點轉基因:與特異突變引入的原理一樣,在同源模版中間加入一個轉基因,這個轉基因在雙鏈DNA斷裂修復過程中會被拷貝到基因組中,從而實現定點轉基因。通過定點轉基因的方法可以把基因插入到人的基因組AAVS1(安全港)位點。這個位點是一個開放位點,支持轉基因長期穩定的表達,破壞這個位點對細胞沒有不良影響,因此被廣泛利用。

CRISPR/Cas9技術三巨頭,認識他們的小夥伴們舉個手讓我瞧瞧~
四、基因沉默gene silencing
基因沉默是指生物體中特定基因因無法表達而失去作用。老規矩,我們先回憶下中心法則:基因要發揮作用,一般來説得從DNA轉錄成RNA,再翻譯成蛋白質。如果説,基因編輯可以通過直接改變DNA序列,實現釜底抽薪式的基因失活操作的話,那麼基因沉默就是發生在RNA的轉錄和蛋白質翻譯這兩種水平上的操作:
前者是由於DNA甲基化、異染色質化以及位置效應等引起的轉錄水平上的基因沉默(tran-scriptional gene silencing,TGS),也就是DNA雖然序列完好,卻無法轉錄成RNA;後者是轉錄後基因沉默(post-transcriptional gene silen-cing,PTGS),即在基因轉錄後的水平上,通過對已經轉錄好的靶標RNA進行特異性降解,使其無法正常翻譯成蛋白,從而使基因沉默。

法國電影《你丫閉嘴》
五、RNA干擾和RNA參與的基因表達調控
RNA干擾(RNA interference, RNAi)是指在進化過程中高度保守的、由雙鏈RNA(double-stranded RNA,dsRNA)誘發的、同源mRNA高效特異性降解的現象【9】。和CRISPR類似,RNAi是細胞對入侵的外源基因的一種自我防禦機制。
研究發現,外源基因隨機整合到宿主細胞基因組內,並利用宿主細胞進行轉錄時,常產生一些特殊的雙鏈RNA(dsRNA)。被整合的宿主細胞能夠識別dsRNA並將它切割成單鏈的小干擾RNA(Small interfering RNA,siRNA),後者被用於形成蛋白-RNA複合物RISC,反過來藉助它本身的序列,去切割降解外源基因轉錄出來用於翻譯蛋白質的信使RNA(mRNA),使得外源基因無法發揮作用,這個過程就稱為RNA干擾(具體機制遠比這裏講的複雜,不再贅述)。
MicroRNA (miRNA) 是另一類由內源基因編碼的長度約為22 個核苷酸的非編碼單鏈RNA分子,它們在動植物中參與轉錄後基因表達調控,每個miRNA可以有多個靶基因,而幾個miRNA也可以調節同一個基因。這種複雜的調節網絡既可以通過一個miRNA來調控多個基因的表達,也可以通過幾個miRNA的組合來精細調控某個基因的表達。據推測,miRNA調節着人類三分之一的基因。就是説,microRNA會影響基因的轉錄和蛋白質的產生。
miRNA是通過什麼方式影響基因轉錄和和蛋白質表達呢?
第一種是切斷靶基因的mRNA分子—miRNA與靶基因完全互補結合,最後切割靶mRNA。是不是非常像RNA干擾呢?科學家認為miRNA可能包含和siRNA類似的作用方式。
第二種是抑制靶基因的翻譯—作用時與靶基因不完全互補結合,進而阻遏翻譯而不影響mRNA的穩定性,這種miRNA是目前發現最多的種類(如線蟲lin-4)。而在植物中極少數的miRNA通過此方式來抑制靶基因。
第三種是結合抑制—具有以上兩種作用模式:當與靶基因互補結合時,直接靶向切割mRNA;當與靶基因不完全結合時,起調節基因表達的作用。
由此可見,microRNA在基因的轉錄和蛋白質表達過程中發揮重要作用。
除了miRNA之外,lncRNA和circRNA也積極參與到細胞的基因表達調控網絡中。lncRNA是長度大於200 個核苷酸的非編碼RNA。研究表明,lncRNA 在劑量補償效應(Dosage compensationeffect)、表觀遺傳調控、細胞週期調控和細胞分化調控等眾多生命活動中發揮重要作用。circRNA則是一類首尾相接的環狀RNA分子,儘管對它的研究起步較晚,但它已經被越來越多的研究證實參與到極其複雜的基因調控系統中,特別是已有研究表明circRNA 在中腦發育、帕金森、阿爾茲海默病和腫瘤發生中發揮重要作用【10,11】。
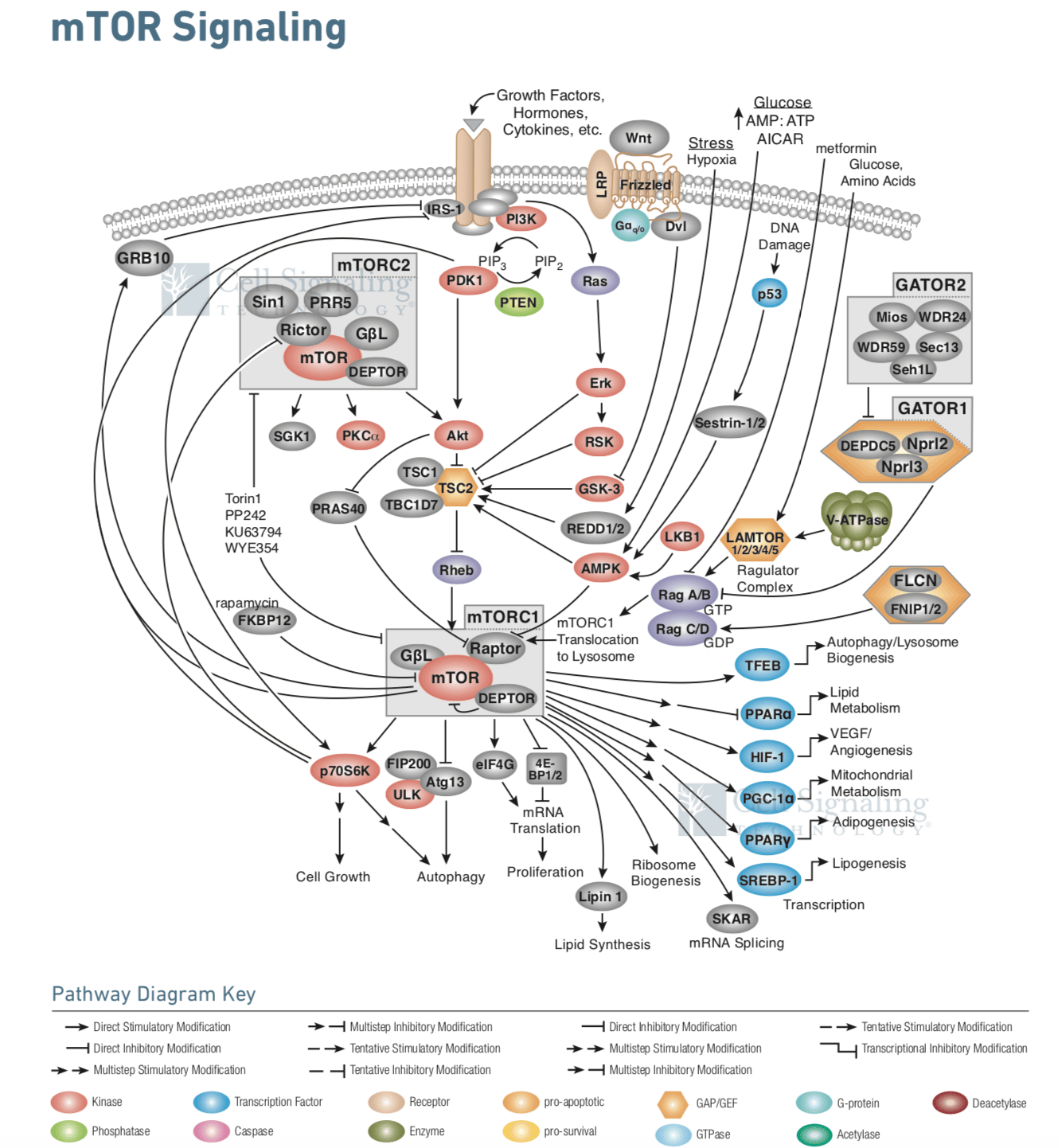
以mTOR基因為中心的細胞信號通路圖示。mTOR是一種主要的生長調節分子,可感受並結合不同的營養因素和環境因素,包括生長因子、能量水平、細胞應激,以及氨基酸【12】
從本文簡單的敍述就可以發現,實際上細胞對於基因功能的控制,也就是基因表達調控,是非常複雜的過程,牽一髮而動全身。上圖所顯示的複雜信號通路,只不過是細胞中複雜調節網絡的冰山一角而已。在實際研究過程中,科學家往往發現,在細胞裏調節了一個基因的表達,就有成千上萬的基因表達受到影響。三個天體在萬有引力下相互作用的規律就已經難以探尋,而數以萬計基因的世界,其複雜的程度恐怕更是超過了大多數人的想象。對基因的理解,我們才剛剛開始。
注
1 雜交過程中同源重組的精確結果不能預知,不等於同源重組是完全隨機的。高中生物知識就告訴我們,同源重組的結果,是等位基因之間的交換,而等位基因的交換結果是有限的幾種。舉個例子,父本在三個基因座上的非同源基因分別是AA,Bb,cc;母本是Aa,bb,Cc。雖然科學家不能預測雜交後這三個基因座上的基因型分別是什麼,但顯而易見A基因座上不會出現B基因同源重組的結果,反之亦然。一定要類比的話,就好像擲六面骰,儘管我不知道下一個骰子會擲出多少點,但終究是一到六點之內的一個整數,絕不會擲出一個無理數來。
2 “…番茄分子育種技術體系的建立很大程度上依賴於人們對控制重要農藝性狀的關鍵基因或 QTL 功能的認知。迄今為止,人們已經克隆或精細定位了 100 餘個番茄農藝性狀相關的基因和 QTL。但目前可通過分子育種技術改良的性狀大多是受單基因控制的質量性狀。對於複雜的農藝性狀(如風味品質和非生物脅迫抗性等),由於缺乏對其分子遺傳基礎的認識,分子育種目前還難有作為。…”
參考文獻
1 修改自百度百科https://baike.baidu.com/item/中心法則/317345
2 Crick, F.H.C. (1958). “On Protein Synthesis”. In F.K. Sanders. Symposia of the Society for Experimental Biology, Number XII: The Biological Replication of Macromolecules. Cambridge University Press. pp. 138–163.
3 Johannsen, W. (1905). Arvelighedslærens elementer (“The Elements of Heredity”. Copenhagen). Rewritten, enlarged and translated into German as Elemente der exakten Erblichkeitslehre (Jena: Gustav Fischer, 1905;
4 美國已種植百萬棵轉基因蘋果樹https://www.guancha.cn/industry-science/2018_09_21_472912.shtml?s=fwckhfbt
5 https://www.sciencedirect.com/topics/medicine-and-dentistry/transgene
6 https://en.wikipedia.org/wiki/Transgene#Ethical_controversy
7 [杜敏敏,周明,鄧磊,李傳友,李常保.番茄分子育種現狀與展望——從基因克隆到品種改良[J].園藝學報,2017,44(03):581-600.](https://www.doc88.com/p-5186377649576.html)
8 Heidenreich M, Zhang F. Applications of CRISPR-Cas systems in neuroscience. Nature reviews Neuroscience. 2016;17(1):36-44. doi:10.1038/nrn.2015.2.
9 https://baike.baidu.com/item/RNA干擾/3743635
10 Liu L, Wang J, Khanabdali R, Kalionis B, Tai X, Xia S. Circular RNAs: Isolation, characterization and their potential role in diseases. RNA Biology. 2017;14(12):1715-1721. doi:10.1080/15476286.2017.1367886.
11 Hanan M, Soreq H, Kadener S. CircRNAs in the brain. RNA Biology. 2017;14(8):1028-1034. doi:10.1080/15476286.2016.1255398.
受水平和時間所限,本文內容必然有不到之處,敬請各位讀者不吝賜教。
本文為觀察者網風聞社區獨家稿件,未經授權,不得轉載。