黑化的打工人:九零後蜜蜂克隆出百萬大軍,差點顛覆南非養蜂業_風聞
返朴-返朴官方账号-关注返朴(ID:fanpu2019),阅读更多!2021-10-19 12:17
More, more, more.
 撰文 | Kestrel
撰文 | Kestrel
蜜蜂(honey bee)是廣為人知的社會性昆蟲,常常被人們用來描述辛勤的勞動者。一個典型的蜂羣由一隻蜂后(也稱為蜂王)、少量的雄蜂和大量的工蜂組成。羣居的蜜蜂在職責上有着嚴格的劃分,蜂后和雄蜂直接參與有性生殖,負責傳宗接代,而工蜂負責育幼、築巢、覓食和防務。(此處響起《打工人之歌》)
為了維持蜂羣在組成上的平衡,蜜蜂的繁殖受到嚴格的調控。其中,工蜂和蜂后是從受精卵孵化而來,均為雌性;而雄蜂是從未受精的卵孵化而來——也就是説,雄蜂沒有爸爸。
通常來説,有性生殖產生的後代同時繼承了父親和母親各貢獻的一套遺傳物質。無論是工蜂、蜂后還是人類,都具有兩套染色體組,也就是“二倍體”(diploid)。不同的是,工蜂和蜂后有16對染色體(總共32條),而人類則具有23對染色體(46條)。而未受精卵孵化而來的雄蜂則只具有正常染色體的一半(總共16條),也就是“單倍體”(haploid,誰讓它沒有爸爸呢)。

不過,在蜂后長時間缺失的條件下,工蜂的卵巢可以激活。但它們仍然不具有交配的能力,只能產下未受精的卵,發育成雄蜂。這種單性生殖的現象也叫孤雌生殖。工蜂單性繁殖的後代都是雄性,因此稱為產雄孤雌生殖(arrhenotokous parthenogenesis,arrhenotoky)。這些雄蜂可以與其它蜂羣的蜂后交配,將攜帶的基因組傳遞下去。
在蜂羣分羣之前,工蜂會在蜂巢內建起4-10個“王台”(queen cell),裏面的幼蟲用蜂王漿餵養,差不多同時完成發育。一般來説,一個種羣只保留一隻蜂后,這些王台中發育出來的蜂后,最強壯的會成為新羣主,其他蜂后都會它被殺死。新的蜂后產生後,就到了分羣(reproductive swarming)的時候。老蜂后會帶着部分工蜂離開,建立新羣。
與此相對應的是,因為一出來就馬上面臨一場生死較量,越早爬出王台對候選的蜂后越有利。需要儘快成熟的蜂后發育是最快的(15-16天),工蜂其次(21天),雄蜂最慢(24天)。
那麼,蜜蜂的性別到底是由什麼來決定的呢?
從分子水平上看,蜜蜂基因組中有一個稱為csd的位點,是控制蜜蜂性別的主要基因位點,全稱是互補性別決定子(complimentary sex determiner)。在蜜蜂種羣中,這個位點的基因大概有19個不同的版本,也就是19個等位基因。雄蜂是單倍體,自然只有一個csd基因。蜂后和工蜂是二倍體,就有兩個csd基因,而且這兩個csd基因的版本總是不同(稱為雜合子),因為版本相同(稱為純合子)的蜜蜂在發育早期就會死亡。
因此,csd位點是決定蜜蜂性別的最上游信號:csd雜合,決定了是雌性;csd純合,蜜蜂無法活下來;只有單個csd基因,決定了是雄性。
也有一些特殊的情況,例如,在高度近交的實驗種羣中,csd位點處於純合狀態的二倍體會發育成雄蜂,但這樣的雄蜂要麼自身不育,要麼交配產生的三倍體後代不育。
工蜂的利他行為可以用親緣選擇學説來解釋。親緣係數(relatedness,簡寫為r)是用來衡量個體間親緣關係的一個量,定義為兩個個體共有某個基因且兩個拷貝都來自同一祖先(例如母子的共有基因來源於母親的父親,即孩子的外公;姊妹的共有基因來源於她們的母親)的概率。拿工蜂來説,產卵的工蜂與其產下的單倍體雄蜂的親緣係數r=0.5,因為雄蜂繼承了其母親一半的遺傳物質,擁有其母親某個基因的概率為1/2。
而工蜂與蜂后產下的雄蜂的親緣係數只有r=0.25,因為兩者都是蜂后的後代:雄蜂擁有蜂后的某個基因的概率是1/2,工蜂作為雄蜂的姐妹,也擁有這個基因的概率也是1/2(還可能是來自另一隻雄蜂),因此兩者共有的這個基因均來自蜂后的概率就是1/4。
但是,蜂羣裏不會只有一隻工蜂產卵,某隻工蜂與所有工蜂產的雄蜂的親緣係數r≈0.125,這樣的話,與只照料蜂后產的雄蜂相比,照料到其它工蜂產的雄蜂反而虧了——為什麼要照料親緣係數更低的雄蜂呢?當然r越高,越是“自己人”。因此,在普通的蜂羣中,工蜂自身不產卵,甚至還會除掉其它工蜂產的卵。
聽起來很損是不是?其實,工蜂的這種行為,是為了增加自身的廣義適合度。所謂適合度,是產生存活後代的能力,而廣義適合度的要求更低:不必自己產生後代,別人的後代包含和自己相同的基因也行。(我的心不禁一顫:多可愛的小生靈啊!)
 圖1 “黑化了,但沒有完全黑化”。腹部黑色比重更大是分辨出海角蜜蜂的一個方便特徵。
圖1 “黑化了,但沒有完全黑化”。腹部黑色比重更大是分辨出海角蜜蜂的一個方便特徵。
這兩個亞種的基因組幾乎完全同源。但是,本文開頭所講到的典型工蜂只可能產下單倍體的雄蜂,而海角蜜蜂卻能獨自產下二倍體卵,發育成工蜂,或者,倘若佔到了王台,發育成蜂后——你沒聽錯,由工蜂產下的蜂后!這種現象稱為產雌孤雌生殖(thelytokous parthenogenesis,thelytoky)。
看到這裏,你可能會想起《生化危機·滅絕》裏面的場景:在愛麗絲和克隆的愛麗絲面前,更多克隆愛麗絲懸浮在營養液裏,被乘放在一望無際的一堆格狀排列的卵形容器中(圖2)。你或許還記得在《生化危機·終章》中,衰老的愛麗西亞將自己的回憶留給了自己的克隆體——主角愛麗絲賦予她完整的“人格”,希望她代替自己生存下去。事實上,澳大利亞悉尼大學昆蟲學家Benjamin Oldroyd也用“轉世”(reincarnation,字面意義是“重新支配肉體”)來指這個無法交配的工蜂產下自己的蜂后克隆的過程。
 圖2 電影《生化危機·滅絕》中相關場景。
圖2 電影《生化危機·滅絕》中相關場景。
的確,過去30年間,數百萬海角蜜蜂克隆大軍曾讓南非養蜂業聞風喪膽。當混在其它亞種的種羣中時,海角蜜蜂是所謂的“社會性寄生物”( social parasite)。這些工蜂不但“違規”產卵、爭搶王台,還散發外激素讓普通工蜂將它們當幼蜂哺育,幾乎享受着蜂后待遇。它們壟斷生殖,致使原種羣漸漸崩潰,然後轉而入侵新的蜂羣。
海角蜂羣還能在缺失蜂后的情況下維持數月,直至產生新的蜂后。上世紀90年代,南非每年有50000蜂羣因其毀於一旦,海角蜜蜂因此被歐美、澳大利亞等地禁運。海角蜜蜂的社會性寄生現象是在巴西引入南非的蜜蜂后首先發現的,幸虧當時處理迅速,不然可能就不叫海角蜜蜂了。
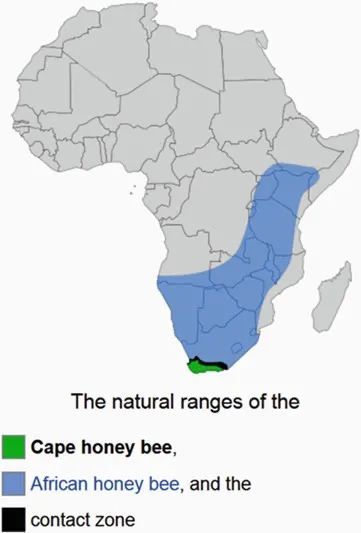 圖3. 海角蜜蜂的地理分佈十分狹小,僅僅侷限在非洲大陸的最南隅(綠色部分)。
圖3. 海角蜜蜂的地理分佈十分狹小,僅僅侷限在非洲大陸的最南隅(綠色部分)。
大家都知道,蜜蜂的身體是黑黃條紋的,但在海角蜜蜂的腹部,黑色的比重更大(圖1),這讓我們可以方便地辨識出它。能自己任命領導(蜂后),自產大軍的海角蜜蜂,可以算是黑化的打工人了吧。
海角蜜蜂的卵也是會經歷減數第二次分裂的,但是染色體分離完成之後,並不會馬上形成核膜和細胞膜。四對染色體,也就是四個原核(pronuclei)排成一列,然後中間的兩個原核(包含的兩套染色體來自非姊妹染色單體)發生融合(“中央融合”),恢復為二倍體。海角蜜蜂的工蜂總是進行孤雌生殖,而蜂后總是進行有性生殖。
 圖4. 一般蜜蜂與海角蜜蜂繁殖模式對比。
圖4. 一般蜜蜂與海角蜜蜂繁殖模式對比。
孤雌生殖是有弊端的,最突出的弊端便是基因組雜合性的缺失,這會降低物種的適應性。減數第一次分裂中期,染色體會發生重組,然後再經歷前面提到的“中央融合”過程。如果某個基因所在的染色體片段與對應的非姊妹染色單體的片段已經交換了一遍,在最終將發育成雌性的卵中,兩套染色體上這個片段將是相同的拷貝。對任一個基因位點,雜合度應該丟失1/3,因為在“中央融合”過程中,選定任一個染色單體,剩下的三個裏面會有一個是和它一模一樣的。
雜合性缺失(loss of heterozygosity)是不利於生存的。一個重要原因是,一些缺陷的隱性基因對應的不利性狀會表現出來。比如在癌症中就經常發現有雜合性缺失,在缺失區域常常發現腫瘤抑制基因亦缺失。研究者曾認為,海角蜜蜂羣體應該會衰退得很快。但是三十餘年過去了,這種情況並沒有發生,這或許是因為海角蜜蜂具有社會性寄生的特性。研究者推測,除了csd基因的純合致死效應,應該還有其它未知的機制在維持海角蜜蜂的基因組雜合度。總之,海角蜜蜂攜帶着許多謎團,還等待着我們去解開。
研究者發現,所有這些海角蜜蜂都是同一只變異工蜂的後代, 這隻工蜂大概生活在1990年。你可能要問,這到底是如何找到的?這裏就要提一下生態學的一種重要研究手段——衞星DNA。
衞星DNA是一類高度重複的基因序列。它是一種分子標記,在生態學中常用來代表更長染色體片段。之所以叫“衞星”,是因為在基因組DNA氯化銫密度梯度離心實驗中,它總是分佈在主條帶以外,好像衞星的位置。
衞星DNA有一類稱為“微衞星”,由2-4個鹼基的序列大量串聯重複而成。分析海角蜜蜂樣本的大量微衞星位點,科學家發現,除了少數突變,所有個體在這些微衞星位點上,要麼是該微衞星序列僅有的兩個版本的雜合子,要麼是這兩個版本其中一個的純合子。這意味着,它們全部來自一隻變異蜜蜂,都在這隻“小祖宗”的孤雌生殖譜系上——堪稱現實世界的Smith。
這聽起來有點驚悚,但你也可以換個角度來理解這個事情。海角蜜蜂變異畢竟是小概率事件,這樣的事件發生過一次的可能性當然要比獨立地發生兩次的可能性大得多(後者概率是前者概率的二次方)。
其實,海角蜜蜂的地理分佈十分狹小,僅僅侷限在非洲大陸的最南隅(圖3)。對於這一區域的養蜂人來説,海角蜜蜂是可愛的存在——好打理,更適應當地的生態環境,更耐受瓦蟎(Varroa destructor,是養蜂業的首要害蟲),在商業價值上完全可以替代普通蜜蜂。而對研究者來説,海角蜜蜂則是最有意思的一個蜜蜂亞種。
參考文獻
[1] Baudry E, Kryger P, Allsopp M et al (2004) Whole-genome scan in thelytokous-laying workers of the cape honeybee (Apis mellifera capensis): central fusion, reduced recombination rates and centromere mapping using half-tetrad analysis. Genetics 167:243–252.
[2] Beye, M., Hasselmann, M., Fondrk, M.K., Page, R.E., Omholt, S.W., 2003. The gene csd is the primary signal for sexual development in the honeybee and encodes an SR-type protein. Cell 114, 419e429.
[3] Jordan Lyndon A, Allsopp Michael H, Oldroyd Benjamin P, Wossler Theresa C and Beekman Madeleine 2008. Cheating honeybee workers produce royal offspring. Proc. R. Soc. B.275345–351
[4] Oldroyd Benjamin P., Yagound Boris, Allsopp Michael H., Holmes Michael J., Buchmann Gabrielle, Zayed Amro and Beekman Madeleine 2021.Adaptive, caste-specific changes to recombination rates in a thelytokous honeybee population. Proc. R. Soc. B.2882021072920210729.
[5] Oldroyd, B., Allsopp, M., Lim, J., & Beekman, M. (2011). A THELYTOKOUS LINEAGE OF SOCIALLY PARASITIC HONEY BEES HAS RETAINED HETEROZYGOSITY DESPITE AT LEAST 10 YEARS OF INBREEDING. Evolution : International Journal of Organic Evolution., 65(3), 860-868.
[6] Rabeling C. Social parasitism. In Encyclopedia of Social Insects (ed. Starr, C.) 836–858 (Springer, Berlin, 2020).
[7] https://entnemdept.ufl.edu/creatures/misc/bees/cape_honey_bee.htm