【CC講壇】張蔚:蝶影重重——揭秘叢林偽裝者_風聞
CC讲坛-CC讲坛官方账号-创新引领未来,传播改变世界。2022-10-21 10:53
大家看到自然界裏存在着各種各樣的動物偽裝者。

偽裝者神秘低調而又生機勃勃,它們身上到底隱藏着自然賦予的什麼秘密?

擬葉偽裝,是一種廣泛存在的動物模擬植物的現象,只要用心觀察,你就能發現這些擬葉動物的蹤跡,有一種因為擬葉絕技久負盛名,它就是枯葉蛺蝶。

我們實驗室主要的研究方向是演化生物學,研究的是非模式生物。
而模式生物,用於揭示生命現象的普遍規律,例如果蠅、擬南芥、小鼠、線蟲等等,它們研究基礎比較好。

然而它們是自然界的滄海一粟,更廣泛存在的其實是非模式生物,具有很多模式生物不具備的一些特徵,可以用來研究更多的科學問題。
一般來講,研究非模式生物相對困難,但是現在科學技術的發展,解鎖了我們深入研究它們的一個可能性,甚至有望將它們建立成研究體系,因此這個研究過程必將充滿挑戰,但是也是我們探索未知的一個必經之路。

我的研究主要關注的就是蝴蝶,關注它們的蝶翅。
蝶翅的花紋是多種多樣的,它們的結構相對簡單,但是它的功能又很複雜,包括有運動、熱調控、求偶、禦敵等等。
這樣一類結構簡單、功能複雜的器官,我們用它來研究一些很有意思的科學問題,例如多樣性如何產生、大自然又是如何塑造這些蝶翅的?
首先隨着我的合作者,西藏的昆蟲學家達娃老師的一個鏡頭,請大家從一個天敵的視角仔細觀察,是否已經發現了獵物?
當鏡頭逐漸拉近,是否又發現了這些蝴蝶擬葉翅花紋的區別?沒有哪兩片葉子是一模一樣的,它展現了多樣的葉形翅花紋。
剛才視頻裏展示的,是枯葉蛺蝶的翅腹面,它們閉合着翅,但是當它們飛翔的時候,就會展示它的翅背面,帶有鮮豔的斑塊。

其實早在1867年,它們就被演化生物學的先驅華萊士所發現,並寫在了書中。
2015年,我在芝加哥大學生態與演化系做博士後研究,有一天,我在讀華萊士的這本書,給我留下了深刻的印象。

枯葉蛺蝶在我國也屬於“三有動物”(有益、有重要經濟價值、有科學研究價值),以保護性的偽裝而出名。
這類蝴蝶的蝶翅有背、腹面迥異的花紋,顯示出它可能受到不同的選擇壓力。
這些假設,還有待於我們進一步地去研究和證明,因此我們這個枯葉蛺蝶的體系,可以用來研究不同選擇壓力下驅動的表型多樣性,此外還可以用來研究背腹面表型不對稱的發育機制。

我們以枯葉蛺蝶的蝶翅為模式研究生物多樣性,可以研究包括遺傳多樣性、物種多樣性,以及受到環境因素和遺傳因素影響的表型多樣性,我就意識到這是一個很好的研究體系。

要研究枯葉蛺蝶,首先還要了解它的生活史。它從受精卵孵化,在幼蟲期要經歷多次蜕皮,然後結蛹,破繭成蝶,這就是一種完全變態發育的昆蟲。
在幼蟲期,蝴蝶離不開賴以為食的寄主植物。因此我們要做蝴蝶的研究,還要去在實驗室飼養它們的寄主植物,給它們做口糧,這對於非模式生物的研究來説,也是一個大工程。
大家可以猜一下,我們把一隻枯葉蛺蝶養大成蝶,需要多少寄主植物?需要十五盆,這也是研究非模式生物的一個特色。
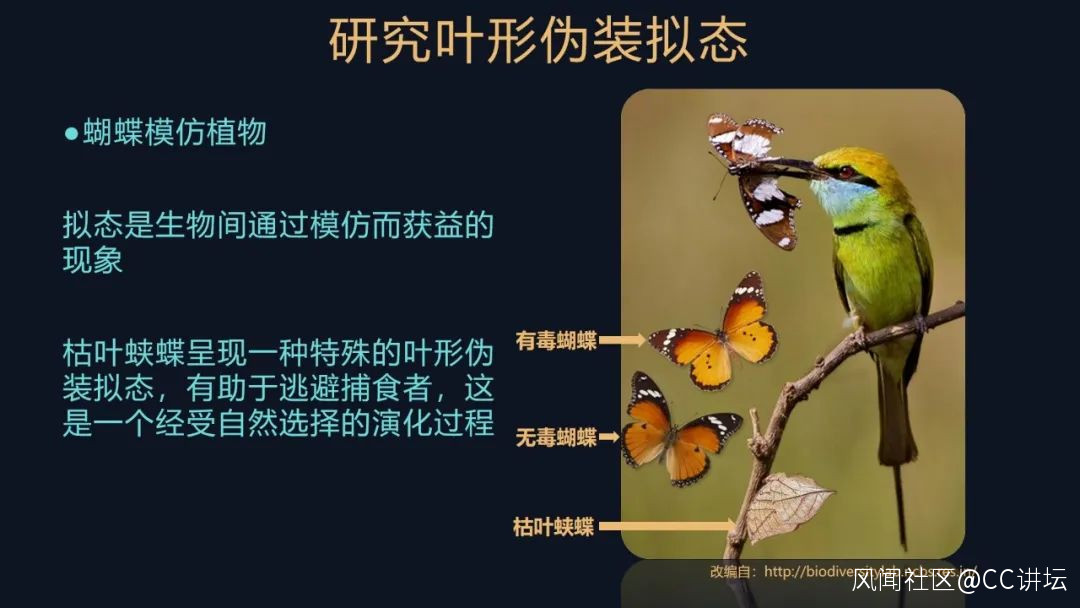
我們今天要介紹的是蝴蝶模仿植物,我們稱之為葉形擬態。
總而言之,擬態的蝴蝶就逃避了捕食者,是有好處的。它們展現的這些巧妙的擬態,其實是經受了大自然的選擇的產物,這是一個被動選擇的演化過程。

生物演化可以追溯至生命起源的一個階段,我們都共享一棵生命之樹,它枝繁葉茂,它也不是一種簡單的線性的演替,讓我們看到的這個世界多姿多彩,這是生物多樣性的一個魅力所在。

演化本身是沒有方向的,也沒有低級和高級之分,但是選擇壓力,在一定的時間可能有一定的方向。
只有選擇壓力本身,沒有可供選擇的材料也不行,所以我們的遺傳信息裏,還會存在一定的變異,這些都是生物演化領域關注的問題。
有了這些研究的框架和手段,我們就做好準備來揭開今天主角的一個神秘面紗。
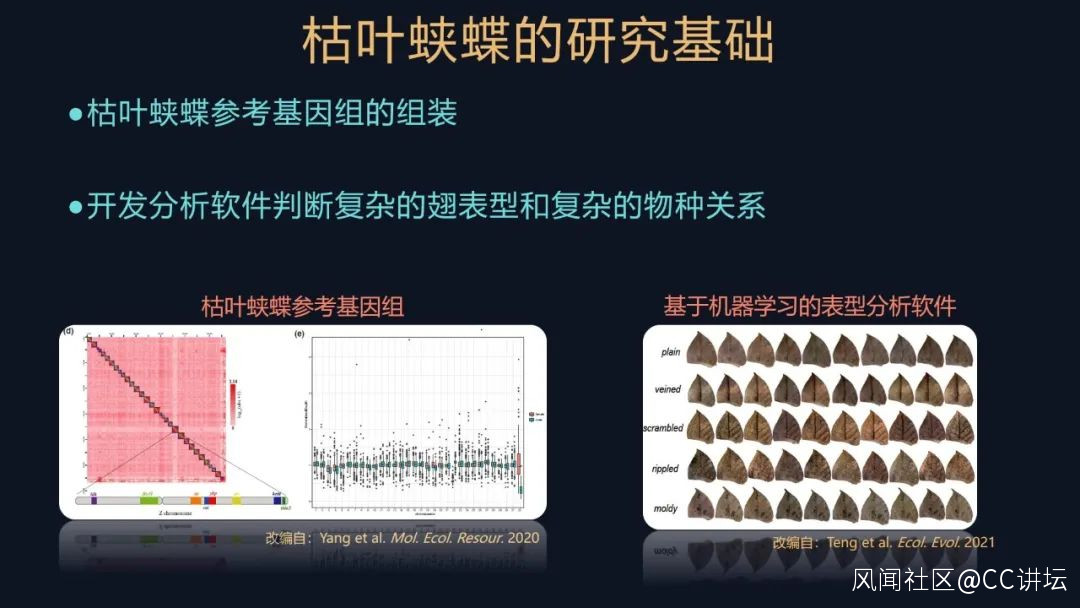
我們首先要參與組裝枯葉蛺蝶中華亞種的參考基因組,我們還實現了在實驗室條件下飼養這些枯葉蛺蝶和開展實驗,此外我們還開發了一個基於深度學習的軟件,來分析它們複雜的物種關係。
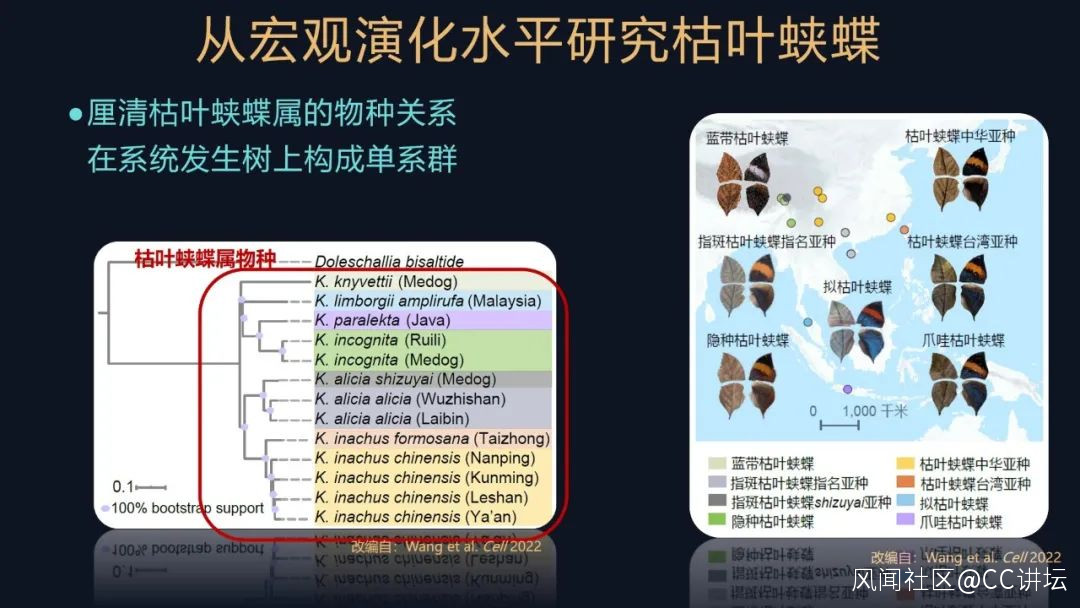
有了這些研究基礎,我們就需要採集樣本了。枯葉蛺蝶在分類學中是一個屬,分佈的範圍很廣泛,還包括多個物種。要弄清楚樣品所屬的物種和它的關係,也是我們開展研究的一個重要的前提。

首先我們從宏觀演化的水平,來解析枯葉蛺蝶屬的物種多樣性,解析其物種的起源和演化。我們就要去到它們自然的生境裏,對它們進行觀察,足跡遍佈典型的分佈的區域。我們採集枯葉蛺蝶的地區,大部分都是熱帶和亞熱帶的雨林,例如有西藏、雲南、海南等等。
我們幸運地躲過了毒蛇,但是很多人沒有躲過螞蝗,沒有躲過“一點紅”的追捕,我們就有博士後曾經被“一點紅”咬了手,手就腫得像個饅頭一樣,但還是堅持每天在收集採集工作。

在這個過程裏最令我印象深刻的一件事,是我們習慣用捕蟲網對它們(蝴蝶)進行採集,有時候難以追上它們隱蔽而迅速的身影,後來我們根據枯葉蛺蝶成蟲喜食一些發酵水果汁液的習性,準備了誘籠,併為它們奉上混合着啤酒和西瓜的一些大餐(我們認為是大餐),但是收效甚微,有時候遇到降雨,把食物衝散了,有時候即使食物完好也收效甚微,吸引不到它們,最後我們恍然大悟,其實雨林裏並不缺乏這些腐爛的植物果實,我們準備的大餐顯然不夠有吸引力。
我們的同學就靈機一動,去當地的一個菜市場買了點啥呢?買了雞腸、雞雜,經過陽光暴曬,可想而知要怎麼樣!它發出刺鼻的味道,讓人避之尤恐不及。但是就是這種味道,特別受蝴蝶的青睞,紛紛現身大快朵頤。
這就太好了!我們終於找到了枯葉蛺蝶的一個大餐,雖然人都被燻得受不了,,但是紛紛地表示這個辦法太絕妙了!
很幸運的是,我們還得到了一些罕見的樣本。這個圖上所示的,就是一些枯葉蛺蝶的,我們主要的採集區域。
我們還釐清了這些枯葉蛺蝶的物種關係,發現它們在系統發生樹上都構成一個單系羣,這些也都是我們後續開展研究的一個基礎。
此外為了研究這些葉形擬態的蝴蝶的起源,我們還收集了很多的蛺蝶科的樣本,包括有一些葉形擬態的其他的蝶,,例如有蠹葉蛺蝶、美洲枯葉蝶等等,聽聽它們的名字都跟葉形、擬葉形有關。
但是就是在這個系統發生樹裏,這些標出了綠色的樣本,可以看出它們構成了一個多系羣。
枯葉蛺蝶屬的樣品是廣泛存在的,一般在一個地理分佈區,只有一個物種或者亞種,我們能採到這些樣品。

但是在一個地方,就是我剛才提到的西藏的墨脱縣,我們獲得了多個物種的樣本,一開始我們以為這是不是弄錯了,沒料到這是個發現,等到我們把樣品的這些基因組全都分析、驗證了以後,我們才相信了這個事實,這就奇怪了,這裏為什麼會有這麼多的枯葉蛺蝶的物種呢?
我們因此就提出了假設,認為這個區域可能是枯葉蛺蝶屬的一個分化中心,或者也有可能是它們在歷史上的冰期的一個避難所?
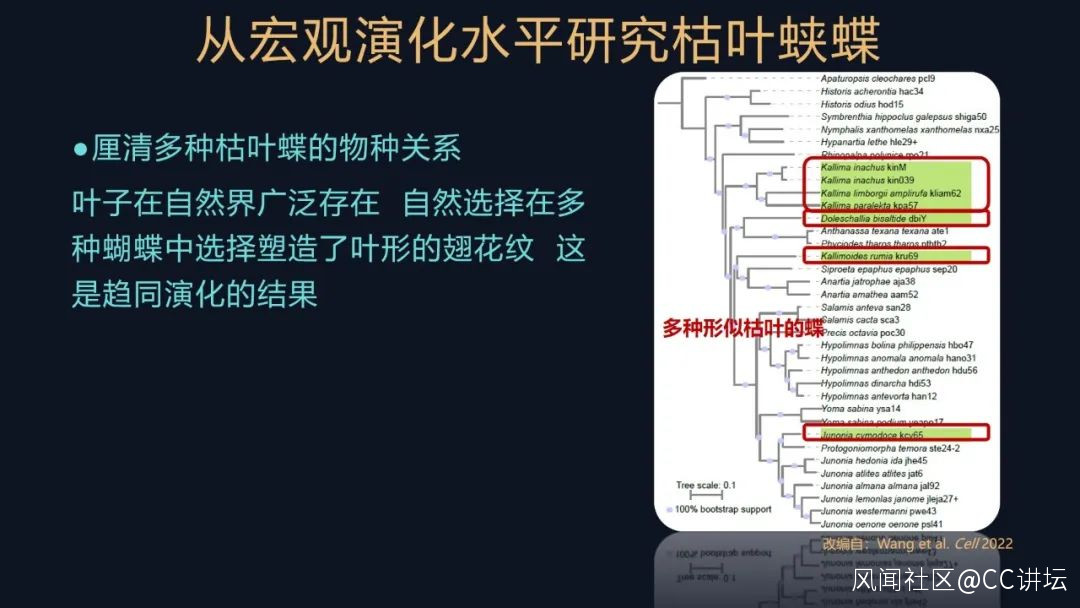
自然界的葉子廣泛存在,自然選擇也多次選擇了這些葉形的表型,這些葉形的翅花紋,就是在蛺蝶科多次獨立的起源,它屬於一種趨同演化。
這樣的話,我們不僅把枯葉蛺蝶屬的物種關係釐清了,我們還把其他擬葉型的蛺蝶科物種的關係都弄明白了,這讓我們感到很開心。
根據我們剛才提出的假設——關於枯葉蛺蝶為什麼在墨脱這個地方有多種物種存在,我們就進一步地研究蛺蝶屬的起源和分化了。
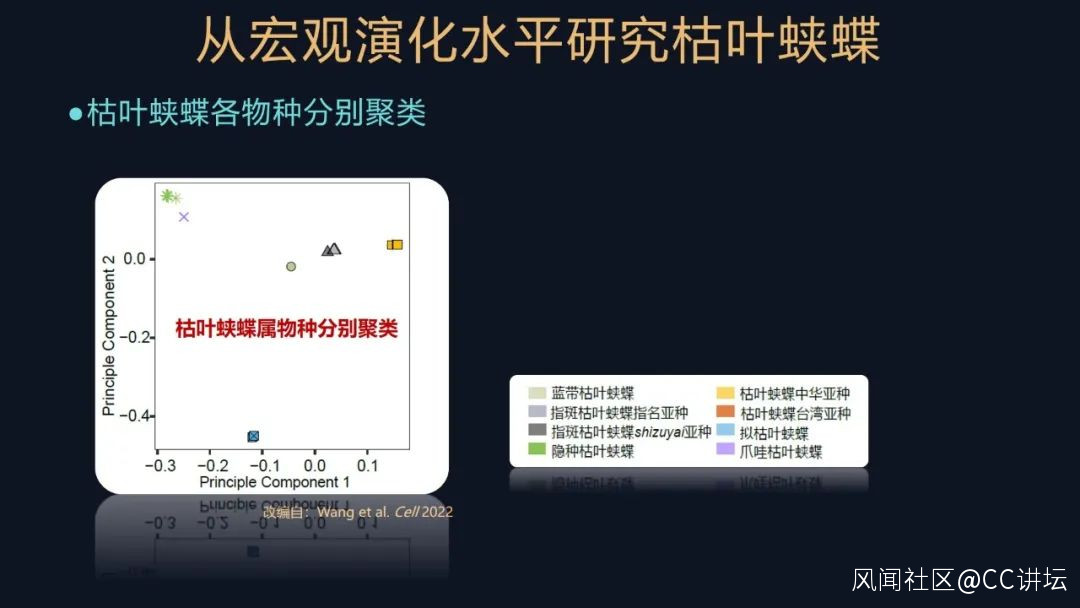
我們對它們的遺傳信息進行聚類和研究,發現這些物種或者亞種,都根據它們的親緣關係聚類了,這説明我們之前對它們的研究和分類還是比較合理和可靠的。
我們又研究了這些蛺蝶物種在歷史上的種羣大小的一個變化,這張圖其實就是根據它們的遺傳信息,反映從現在到以前,這些不同物種種羣的一個變化情況。可以看到大部分的枯葉蛺蝶屬的物種或亞種,有比較大的種羣,但是都是在末次冰期這種歷史時期,它經歷了一個瓶頸效應的影響,也就是它的種羣有收縮、有波動,但是總體來説在喜馬拉雅東部地區的這些物種,都顯示出了比較大的種羣,再次説明這個區域對於枯葉蛺蝶屬是比較重要的。
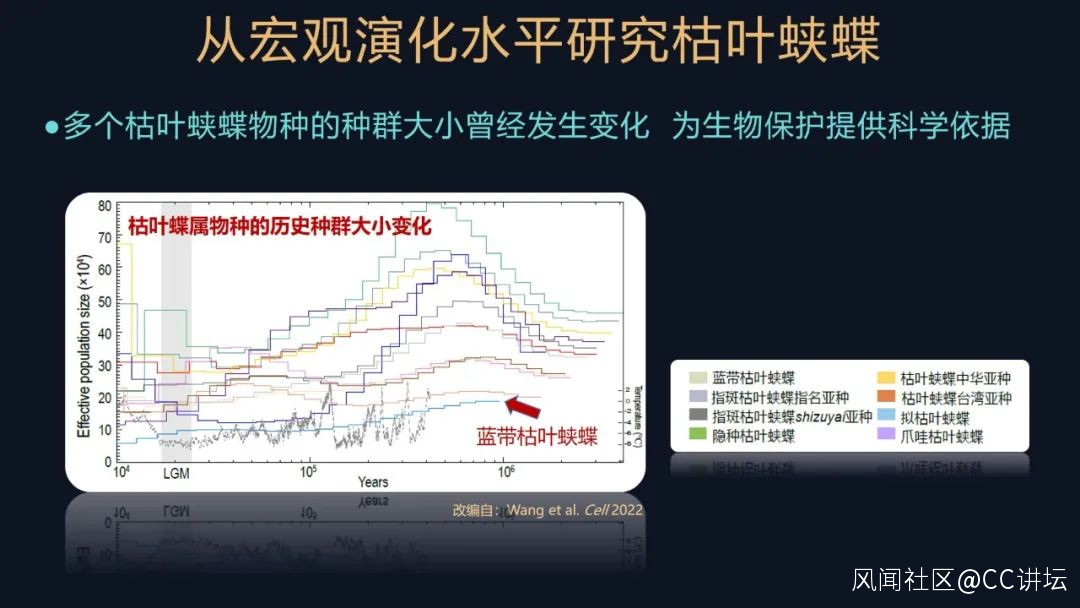
還有一個例外,就是這張圖上的藍帶枯葉蛺蝶,它始終維持着一個比較小的種羣,其實這個蝴蝶目前在我們國家,僅僅在墨脱縣發現分佈了,它的樣品非常難以獲得,我們大概要採集幾年才能獲得這樣一個樣本,再次印證了它的種羣比較小的這一點。
我們其實做昆蟲研究,很難像研究大型的動物一樣,對這些昆蟲進行抽樣計數,你仔細地把它數一數,這是不可能的,但是我們通過基因組學的研究手段,可以獲得它們這些種羣的信息,這也為對它們進行生物保護提供了很重要的科學依據。
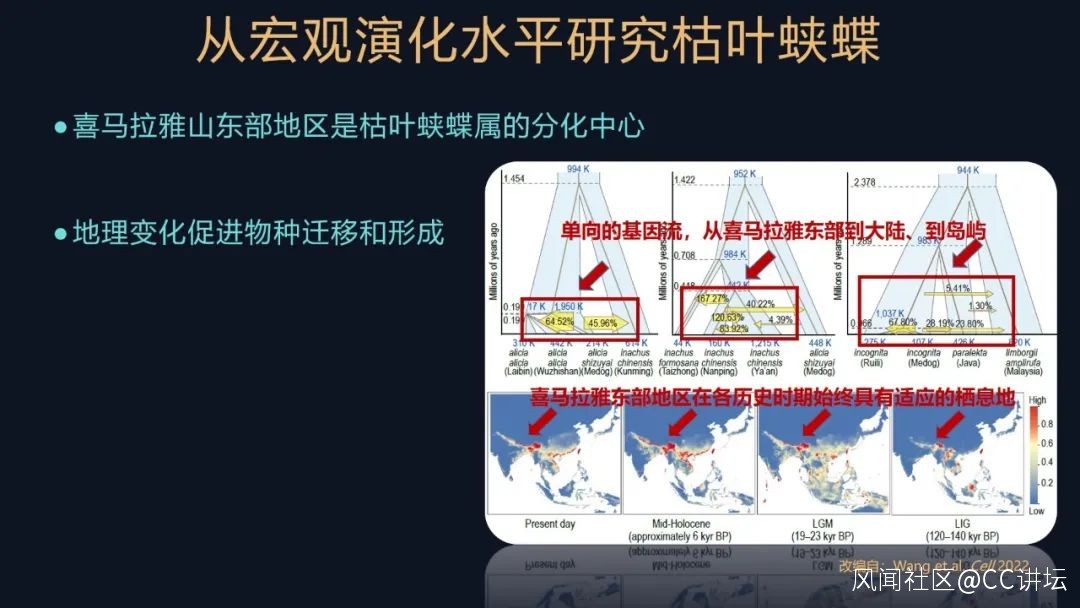
我們還進行了一些基於模型的分析,研究不同的亞種,或者是不同的種羣之間的基因流。所謂基因流,就是指遺傳信息的一個交換。
我們這個研究就發現了枯葉蛺蝶屬都有一些單向的基因流,方向是什麼樣的呢?例如從喜馬拉雅東部地區向大陸地區流動,或者是從大陸地區到島嶼地區流動,反映了枯葉蛺蝶的一個遷移和分化的過程。這個過程是需要非常長的歷史時間的,非常壯麗的一個遷移的過程。
我們還對它們歷史上適宜的棲息地進行分析,發現喜馬拉雅東部區域始終有它們適宜的一個生境,而冰期由於海平面下降,會產生一些陸橋的結構,又會幫助這些蝴蝶向島嶼進行進發,進而分化形成了一些島嶼的物種,這就是一幅波瀾壯闊的遷移圖景,持續數十萬年。
枯葉蛺蝶屬的分化時間,和青藏高原隆起的時間是相符合的。高原隆起產生的海拔落差,會形成多樣的生境,它有助於孕育新的枯葉蛺蝶的物種,到現在為止我們就揭示了枯葉蛺蝶屬的演化和起源的歷史。
它們的葉形翅型又如何產生呢?我們將從微觀演化的水平解析它們葉形表型的一些遺傳機制。

我們先以其中的一種,枯葉蛺蝶的中華亞種作為一個模式,對它們進行觀察,建立一些家系。我們把具有不同葉形表型的這些個體進行組合,看它後代會有怎樣的表型。
在這個過程裏,我們發現着實不少,它一共有十種離散的表型。

有比較樸素,沒有花紋的,我們命名為無主脈表型;還有的是有一條深色的主脈,我們命名為有主脈;有的還有側脈,有分支了;還有的有黴斑、有水波紋;不同的表型還會有一定的規律進行組合。這其實是非常經典的遺傳學的研究方法,只不過我們把它用在了非模式的生物上了。
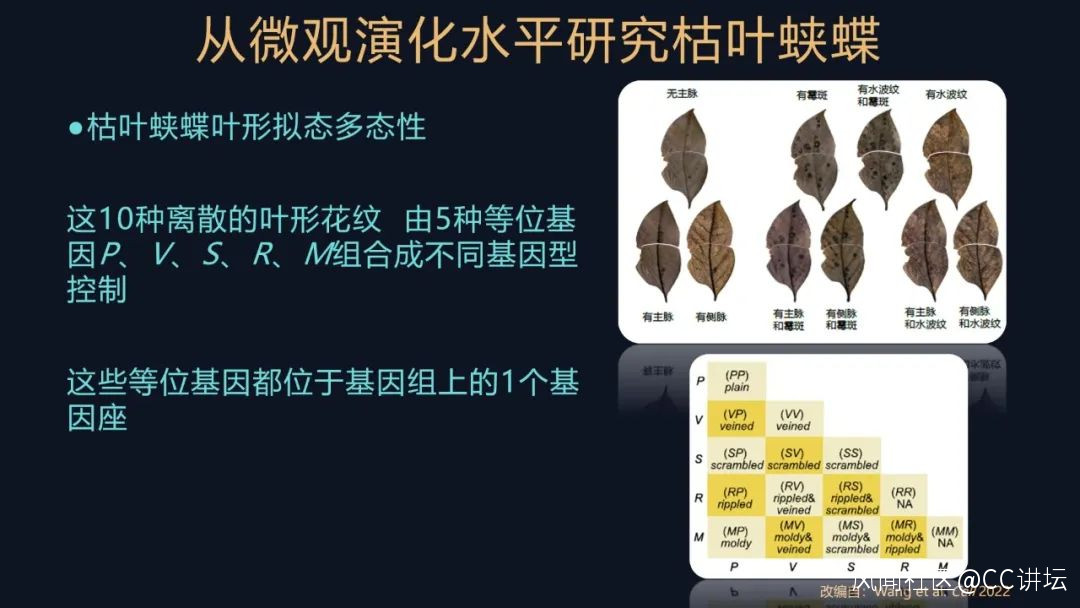
我們還對這些表型進行一個判斷,推測這十種表型,由五種等位基因控制,都位於一個單基因座,它們通過組合形成不同的基因型,進而控制這十種表型,但這些都是基於我們初步的實驗結果提出的假設,我們還要對它進行驗證。
但是逐漸我們的實驗脈絡就愈加清晰了,發現了這麼多的葉形表型 非常地驚喜,但是也意味着整個的研究過程比我們預期的還要複雜。
但科學研究中最有趣的事兒就是在此,莫過於你提出一個或回答一個問題的過程裏,陸續發現更多有意思的和待解答的問題。
我們用飼養獲得的個體,去根據它們的不同的表型進行分組,然後對它們進行全基因組的關聯分析,解析控制葉形擬態的遺傳機制。
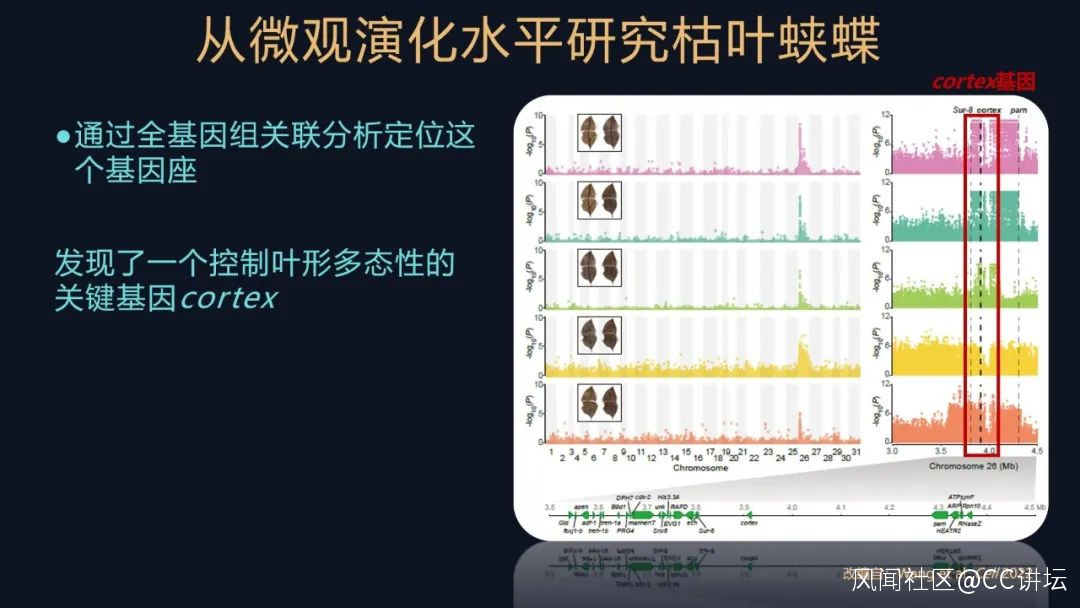
我們把兩兩表型進行一個分析比較,都在基因組的同一個位置發現了類似的信號,這個信號裏有啥呢?
我們發現了一個控制擬態的關鍵基因,它叫做cortex
這個發現又是一個意外之喜,為什麼這麼説?cortex這個基因,它是一個大名鼎鼎的基因。蝴蝶的發育過程相對來講其實是比較保守的,有一些工具盒基因各司其職,它們負責起到一些重要的作用,cortex基因就是其中的之一。

它已經被報道參與了多種受到自然選擇產生表型多樣性的例子裏了。

這個基因的缺少確實會使枯葉蛺蝶的翅的腹背面花紋受到影響。進一步我們還想知道一下,這個多樣的葉形表型,它是如何維持的?
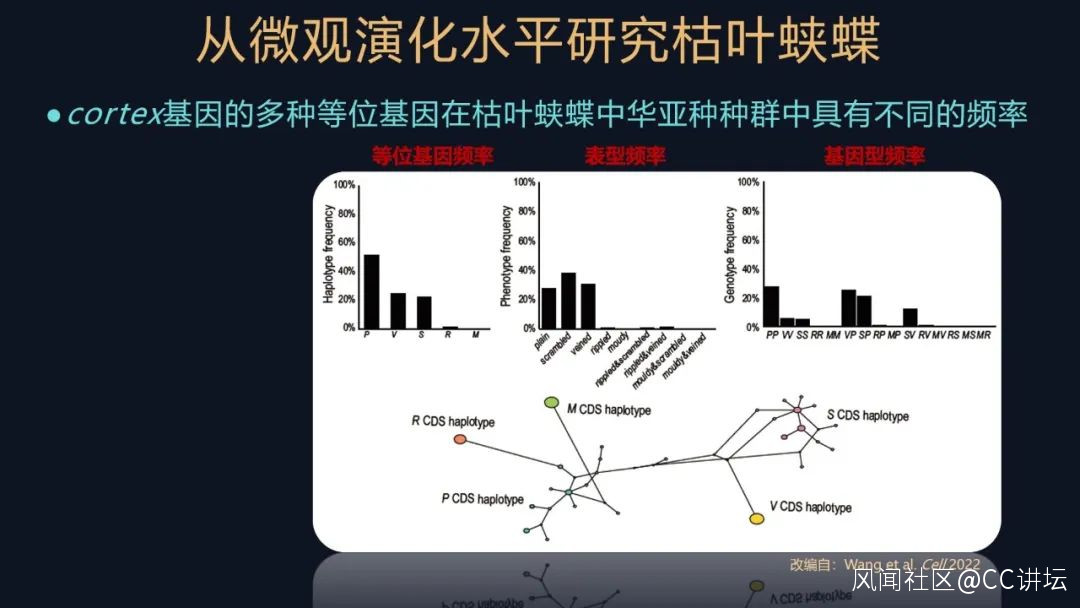
我們就去統計枯葉蛺蝶中華亞種不同的基因型和表型的一個頻率,這又是一個非常經典的遺傳學的研究方法。發現它們都各不相同,説明cortex不同的等位基因,它們已經演化了比較久的時間了。其實不僅僅是在枯葉蛺蝶中華亞種的種羣,我們在很多的枯葉蛺蝶屬的物種,都發現了葉形擬態的多態性。
這個結果讓人眼前一亮,又是一個未知忽然出現的驚喜。
這可能是一個平衡選擇的例子,這些等位基因產生的時間,剛好跟整個枯葉蛺蝶屬的物種分化的時間是能對得上的,進而又跟一個什麼時間對得上?就是跟青藏高原隆起的時間又對上了。這是一個受到自然選擇維持的一個長期的平衡選擇的一個例子,它很罕見。

為什麼説罕見呢?要維持長期的平衡選擇,對於這些物種來講壓力其實是比較大的。
我們建立了模型證明,它受到了一種負頻率依賴的選擇,和一種純和的劣勢。
例如在枯葉蛺蝶中華亞種裏,我們發現有一些表型,它只能以雜合的形式存在,就是有一些剛剛產生出來的,例如黴斑表型,沒有一個純和的狀態,這説明什麼?可能它還是新產生出來的等位基因,還有一些劣勢需要通過雜合狀態來彌補;
而負頻率依賴的選擇又是什麼意思?説明什麼表型多了、頻率多了,它反而不好。試想如果捕食者看到了整齊劃一,千篇一律的葉形,比我們眼中的假花、假草還不能再假了,一眼假,很快就會被捕食者所識破。
所以它們的這種多種多樣的葉形,恰恰好是自然的偽裝,所以這是自然選擇的一個結果,也是枯葉蛺蝶屬的一個生存策略。

我們在微觀和宏觀演化水平研究了枯葉蛺蝶屬的起源和多樣化的工作,最近也以封面文章的形式發表在了《細胞》雜誌上。

我們究竟從研究枯葉蛺蝶的葉形偽裝裏獲得一些怎樣的啓發?
我們首先解鎖了一個新的研究體系,通過它還研究了生物多樣性的演化和遺傳機制。
關於葉形偽裝擬態的研究也都是剛剛開始,這是枯葉蛺蝶作為研究體系的一個魅力所在,也是科學研究的魅力所在。

孤舉者難起,眾行者易趨,當我們克服重重困難,再次回顧的時候,非模式生物的研究,它們就都變成了科學求索道路上的風景。