COVID船新版本前瞻_風聞
Nekromanzer-Ʞ!lןj0ʎ, Çʎņ!ȼḁĻ01-13 15:19
從未知原因的電子煙病毒開始算的話,現在已經是新冠流行的第四個年頭了,離人類走出疫情似乎越來越遠。
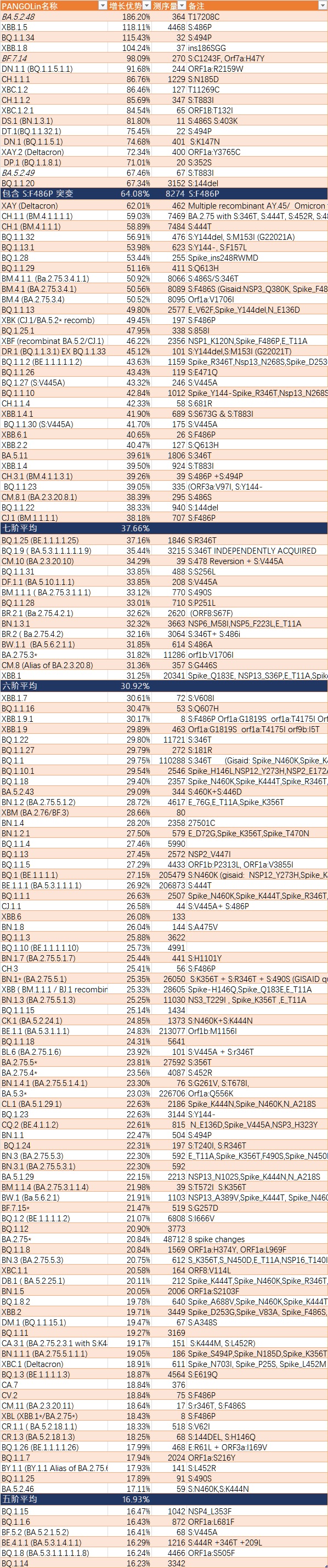
新的天梯榜,可以看到幾個國內流行的變種BF.7.14,BA.5.48,BA.5.49
由於一時半會還沒有競爭對手所以有了不符合自身實力的排名
總結一下上一年的COVID19的演化史,病毒學家們提出了“變種湯”(Covid Variant Soup)這樣的趨同演化(Convergent Evolution)的概念,由於US CDC的刻意淡化(Just common cold/Mild)以及WHO的不作為,人們不得不把一整鍋湯都叫做“奧米克戎”,而這鍋湯裏含有677個奧米克戎變種以及89個奧米克戎重組株變種(每天都在迅速增加),它們的性質各不相同,偏偏要被當作同一種東西來討論,彷彿它們的傳播能力、致病性等等都是一塵不變的,極大地誤導了公眾。
既不神聖,也不羅馬,更非帝國的神羅甚至不如桶裝水
把一大鍋變種都叫做奧米克戎來討論就是這樣的謬誤
有人説了,難道這鍋湯不都是B.1.1.529的分支嗎,問題是WHO有過給同一祖系的不同分支給與不同希臘字母編號的前例,比如Gamma、Zeta、Theta屬於一根藤上(B.1.1.28)的三隻瓜,分別都得到了希臘字母編號;此外Kappa和Delta也是同一根藤(B.1.617)上的不同分支,也都同樣得到了不同的希臘字母編號。
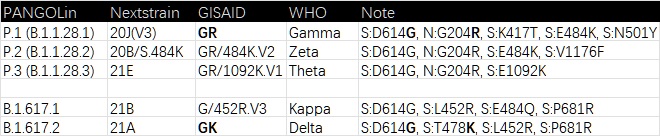
為什麼同樣是B.1.1.529這跟藤上出來的五兄弟,就沒有這樣的待遇了?除了老三(BA.3)以外哪個不是大殺四方,而BA.1、BA.2、BA.5更是全球制霸,這三兄弟不比起其它有WHO編制的強嗎(Delta除外)?
新冠可不管這屆不行的裸猿是不是唯心主義,WHO這樣的國際組織是不是搞歧視性政策,輕視它的存在,它就會慢慢發育直到搞出大新聞,直到讓疫苗、單抗紛紛失效。
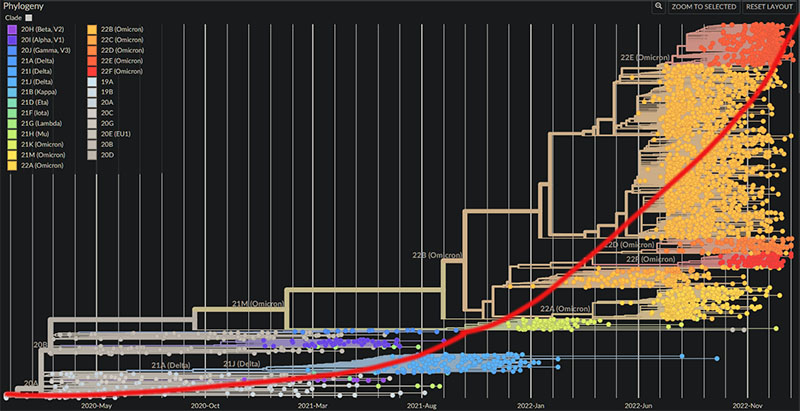
矩狀系統發生樹,可用看到COVID19的演化速度在加快
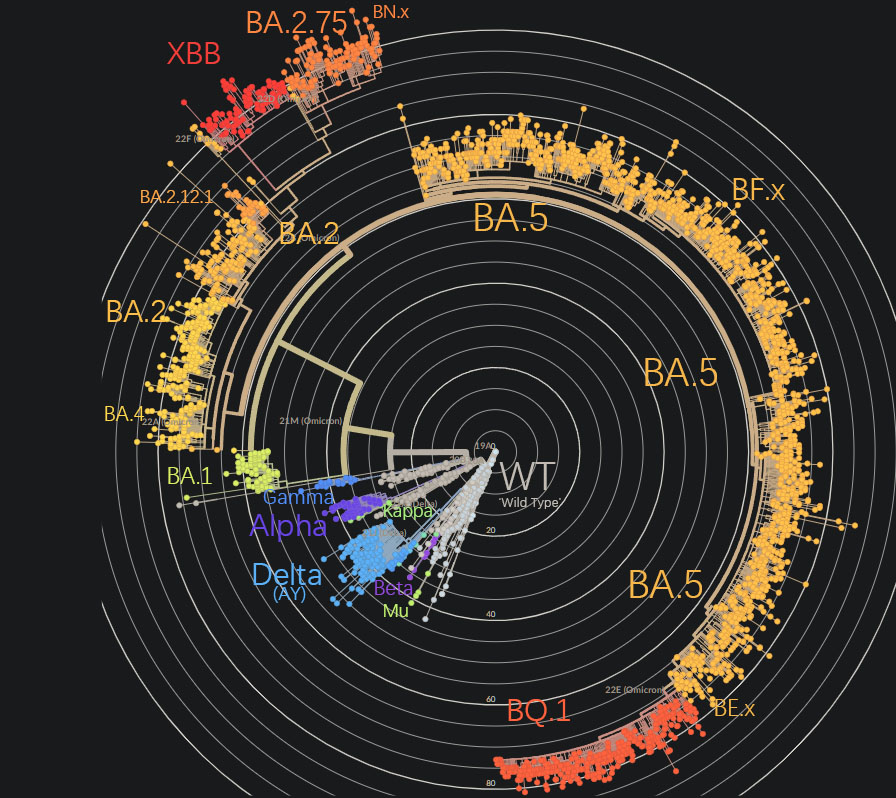
換做放射狀系統發生樹,可以看到所謂的Omicron是個多麼龐大的譜系
尤其是BA.5產生了眾多的分支
現在的COVID19已經分化到了離譜的程度,拿最新的XBB來説,它的抗原距離已經大到難以中和的程度了,就這樣仍然要被稱作奧米克戎,絲毫不管這樣的討論是否還有價值。
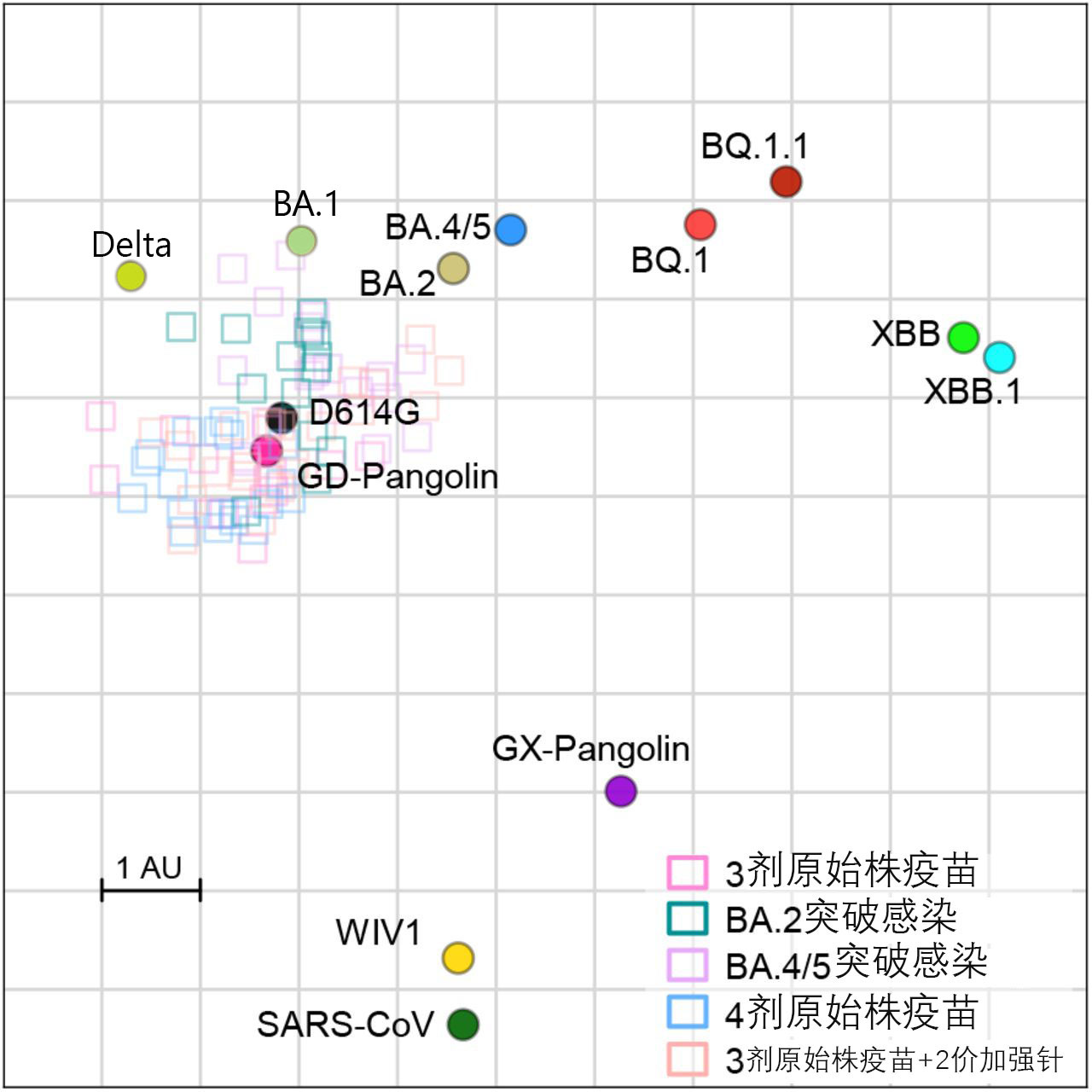
根據 doi 10.1016/j.cell.2022.12.018 Figure S3
以及 10.1126/sciimmunol.abq4450 Fig. 4 修改
上圖是由中和血清數據製作的新冠變種與相近的嚴重急性呼吸道綜合徵相關β冠狀病毒(SARSr-Betacoronavirus, sarbecovirus**)的抗原圖譜,方框表示中和血清位置,座標軸為抗原距離,任易方向上的一個抗原距離單位(Antigenic distance unit, AU)對應於中和抗體ID50滴度的兩倍變化。D614G為B.1株的標誌性突變,WIV1為Bat SL-CoV-WIV1的簡寫,是宿主為蝙蝠的SARS-CoV-1的原始株;被懷疑是SARS-CoV-2中間宿主的穿山甲攜帶的是Pangolin SARSr-CoV-GD,但是它離COVID19野生型(Wild Type,WT)的測序相似度還不如Bat BANAL-52**。
部分裸猿鄉愿地奢望與新冠共存,只是看了這麼多隨後發生的越來越離譜的症狀,以及它會造成免疫系統的紊亂由此導致的與其它疾病共感染的案例,不禁要問一句:但是代價是什麼?
趨同演化/共循環
在COVID19進入第三年的版本之前,不同變種之間是競爭的模式,更具傳播優勢的毒株會把其它的變異株甩在身後,最後養出的蠱王是兼具傳播性和致病力的Delta,Lambda和Mu。不過等到從其它物種回傳裸猿的Omicron出現以後,大多都被拍死在了沙灘上。
在進入船新的版本之前,COVID19趨同演化的現象其實就已經出現了,當時在全球的很多地方,在毫無關聯的病例中(尤其是Delta的子株AY系列)出現了大量的趨同變異,只不過後來的奧米克戎BA.1海嘯把大家的注意力都吸走了。等到BA.2的浪潮似乎就要過去後,人們發現全球各地出現了BA.2、BA.4以及BA.5家族共同爭奪份額的情況。
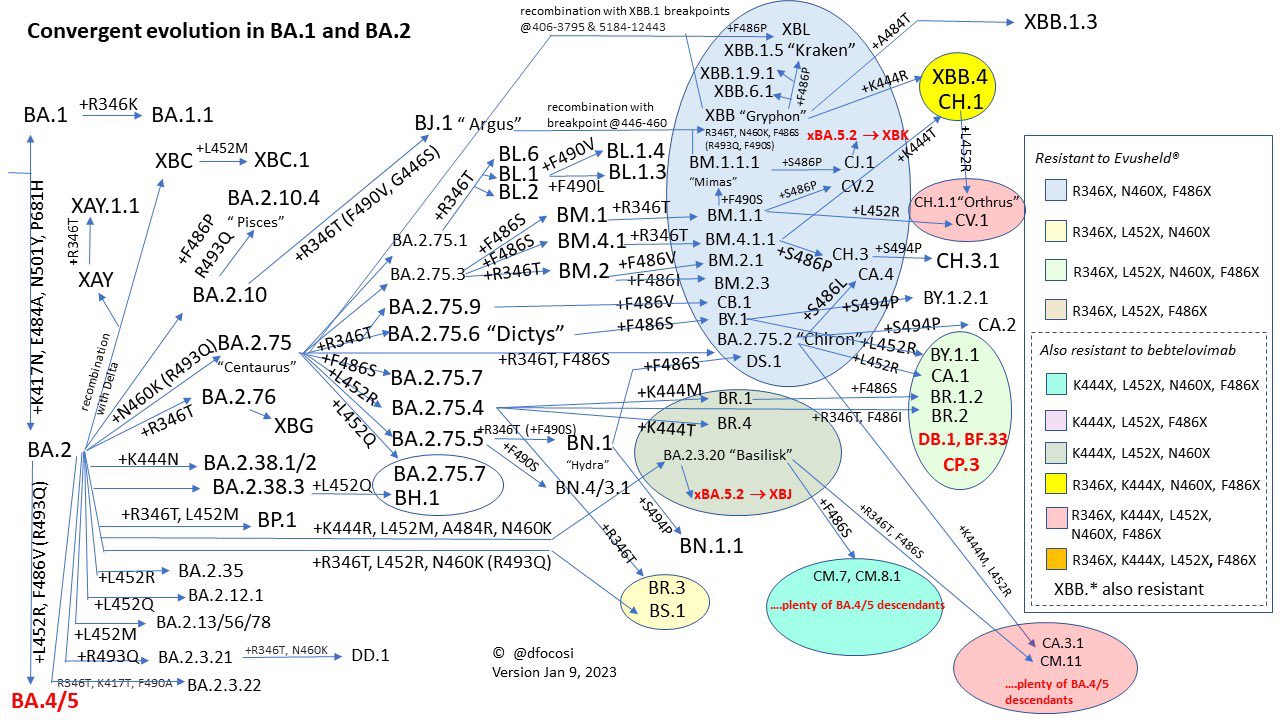
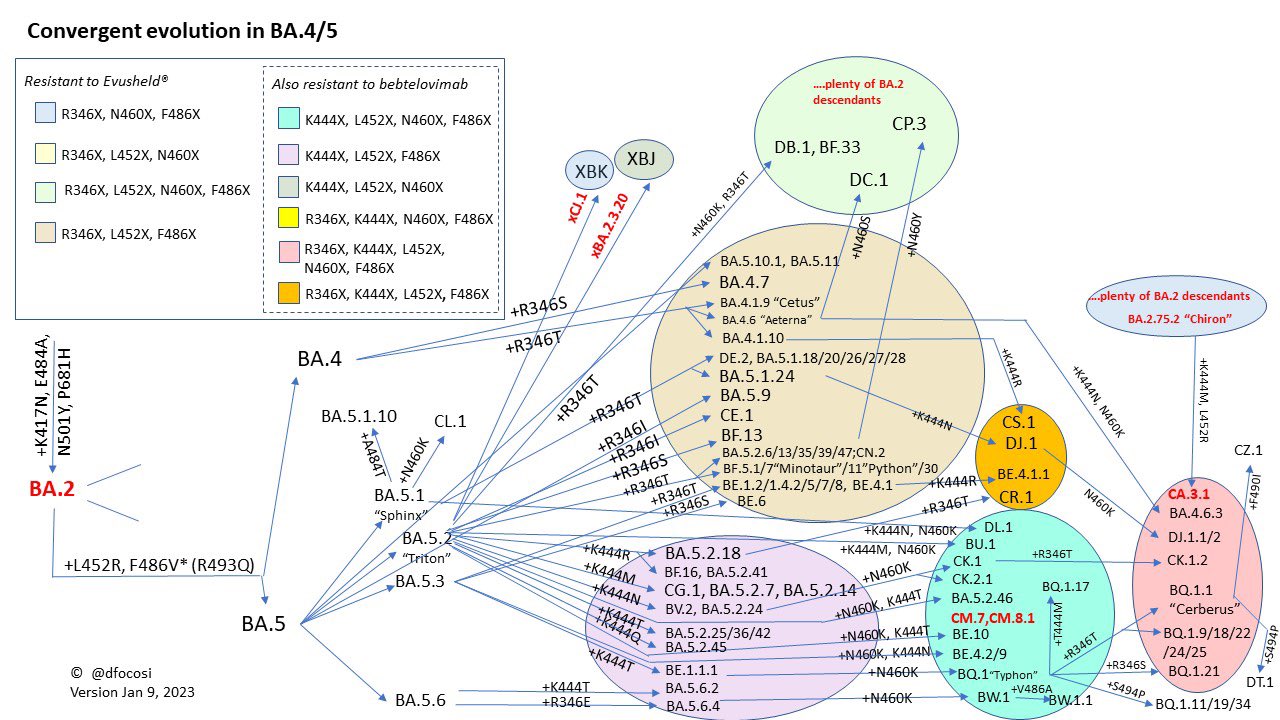
Daniele Focosi教授製作的Omicron“趨同演化”圖
這段時間人們發現Omicron的各個分支都在朝着免疫逃逸的方向瘋狂演化,於是就有了“天階序列”,根據計算病毒學(Computational Virology)大佬Jesse Bloom的分析,將刺突蛋白的受體結合域(Receptor-Binding Domains, RBD)中那些熱門的突變位點做了一個總結,只要突變出現以下位點:R346,K356,K444,V445,G446,N450,L452,N460,F486,F490,Q493,S494,每獲得任何一個這些位點的突變則稱為“一階”。
目前這套模型最高只有12階,但這並不代表COVID19就只有這麼點免疫逃逸的潛力,因為RBD僅僅是刺突蛋白(共1273個位點)的結構上的一小段(位點319~541),除此之外還有更多其它結構上的突變位點並沒有算入在內,比如同樣是刺突蛋白中的氨基端結構域(N-Terminal Domain,NTD,位點14~305)的Y144Δ等。
根據不同變種的“階位”可以大致估算出它們的相對增長優勢,階位越高的變種流行起來的可能也越大,這段時間內具有這樣特點的大概有BF、BJ、BN、BQ、BR、CH。
與此同時,COVID19的另一種變異方式開始大量出現,就是重組,被BA.2壓制的BA.1和被BA.1壓制的Delta(AY)藉此實現了穢土轉生。重組株的出現標誌着宿主可能被不同的毒株同時感染,同樣意味着這幾種毒株直接的交叉免疫幾乎不存在了,靠着疫苗+突破感染試圖獲得更高的中和抗體的邪路(至少到Delta時期如果身體能扛得住的話這個歪路子確實有點效果)至此已經走不通了。
一代毒王的誕生
在南亞次大陸這個神奇的地方,不得不説新冠可能真不是什麼太嚴重的問題,面對那邊還未解決的甲類傳染病,新冠可能真的是Mild,也就意味着那裏無論養出什麼毒株都不要奇怪。
6月初,印度開始上傳一些躍遷式突變的BA.2子系,其中包括BA.2.74、BA.2.75和BA.2.76,根據曹老師的分析BA.2.75的hACE2受體結合親和度(hACE2 binding affinity)非常高,意味着它在宿主體內發生突變的能力又增強了。
BA.2.75比起BA.2增加了刺突蛋白上的K147E, W152R, F157L, I210V, G257S, D339H, G446S, N460K, R493Q這些突變。它的一個子系BA.2.75.3又獲得了S:R346T。然後在它的後代中又得到了S:F490S,也就是BM.1.1.1。
幾乎是與此同時,BA.2.75的一些子系和BA.2.10的子系BJ.1裏都出現了一些帶S:F486P的變異,此時有關於486位點的探討還僅限於免疫逃逸,這個F486P比起F486S來説要損失一些免疫逃逸能力,當時並未能引起足夠的重視。
不久後在新加坡發現了一個BM.1.1.1和BJ.1的重組,據推測它形成於南亞次大陸,發現時離它在新加坡肆虐已經不遠了。這個重組株被PANGO委員會命名為XBB。
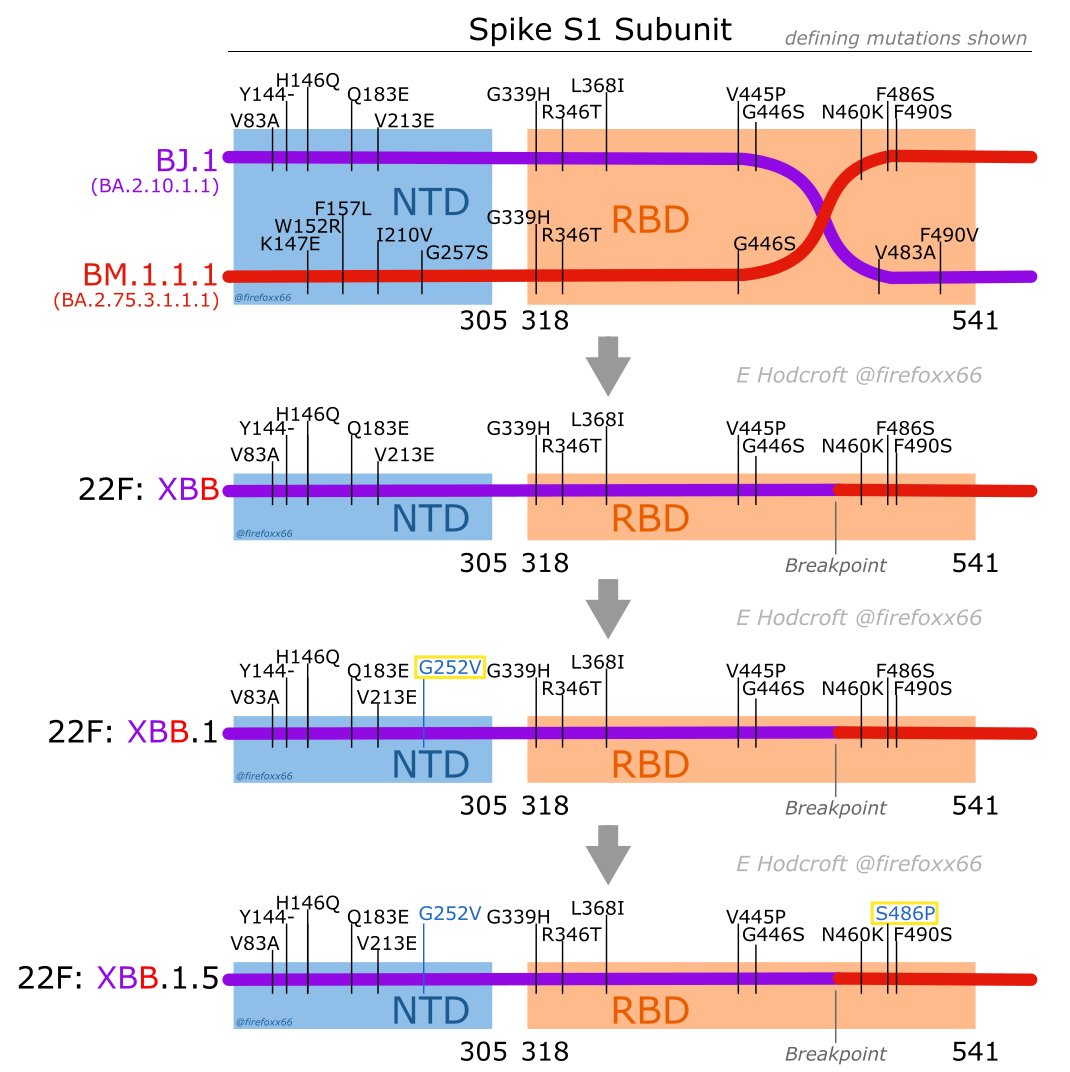
圖為Emma Hodcroft老師對XBB重組和繼續演化的示意
可以看到雖然BA.2.75的子系和BA.2.10的子系雖然都有一些出現過S:F486P,但是在講究免疫逃逸的版本里它並不是版本答案,所以重組的XBB得到的是S:F486S。
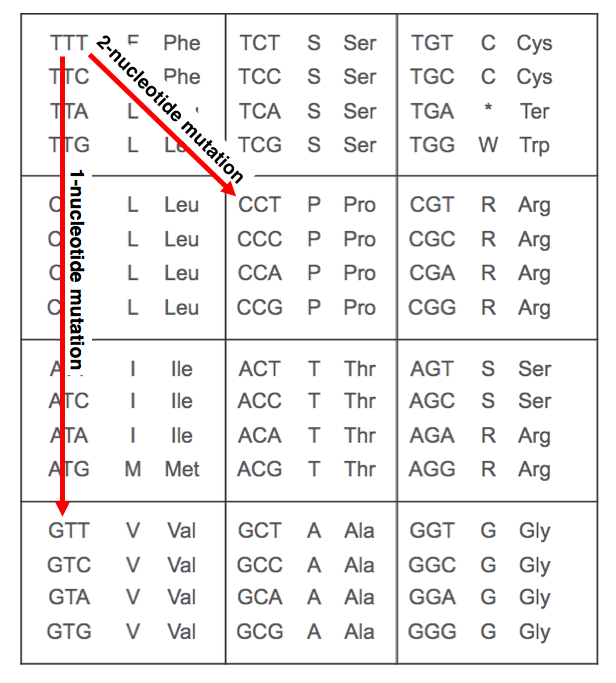
核苷酸與氨基酸的編碼關係,來自Ryan Hisner老師的講解
而從S:F486S到S:F486P需要1次核苷酸的改變(T23018C,TCT→CCT)。這樣的變化在紐約最終得到了完成,於是完全體的XBB.1.5誕生了。
相關內容請見:
船新的版本
XBB.1.5橫空出世以後經過了一小段的蟄伏(有近兩週未被測序,要麼是發生了隱秘傳播,要麼就是CDC的疏忽)後,開始突然爆發。此時的北美尚處於BQ和XBB家族的你爭我奪當中,帶有S:F486P的XBB.1.5打破了平衡。這時候人們開始意識到“受體結合親和力”這個此前並未受到太多重視的特性有多麼重要了。
在F486的19種可能的突變中,P有着最高的hACE2受體結合親和力,較高的RBD表達(RBD Expression),不弱的免疫逃逸能力,如果不考慮變異的難度,恐怕是這個位點上最佳(對於COVID19而言)的變異選擇。
XBB.1.5在攢夠免疫逃逸的階位以後,以損失一點免疫逃逸能力的代價換取了受體結合親和力的高額回報,這恐怕意味着毒株之間的軍備競賽要進入下一個版本了。
現在不少高階位的毒株基本上都帶有S:F486P,S:F486I或者S:F486V,這幾種突變的受體結合親和力依次降低。BQ家族的問題在於BA.5家族的底子就是S:F486V,當時只用了一個核苷酸的變化就完成了S:F486V(T23018G,TTT→GTT),它的免疫逃逸能力最強,但是受體結合親和力就差了一些,想要再變成S:F486P至少還要兩次核苷酸的變化(G23018C以及T23019C,GTT→CCT),而且據推測這樣的變化難度還不小,如果BQ家族不能在其它方式上動點子的話很可能要被XBB.1.5給反推。
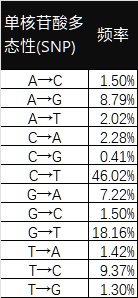
複習下高中(?)知識
目前稍微領先的BQ家族可能不得不走一條“艱難的路”
除了S:F486P以外,現在不少的毒株開始在S2亞基甚至開放閲讀框架(Open Reading Frame, ORF)、核衣殼蛋白 (Nucleocapsid Protein, N)以及非結構蛋白(Non-Structure Protein, NSP)上搞事情了,後面這些區域的相關研究跟S蛋白比起來要少得多,尤其是NSP基本抓瞎,只能猜測這些很多都是跟致病性相關的,換人話説,就像瘟疫公司(Plague Inc.)裏預測的那樣,點完了傳播力就該點致病性了。聽上去就不是什麼好消息。
種子選手
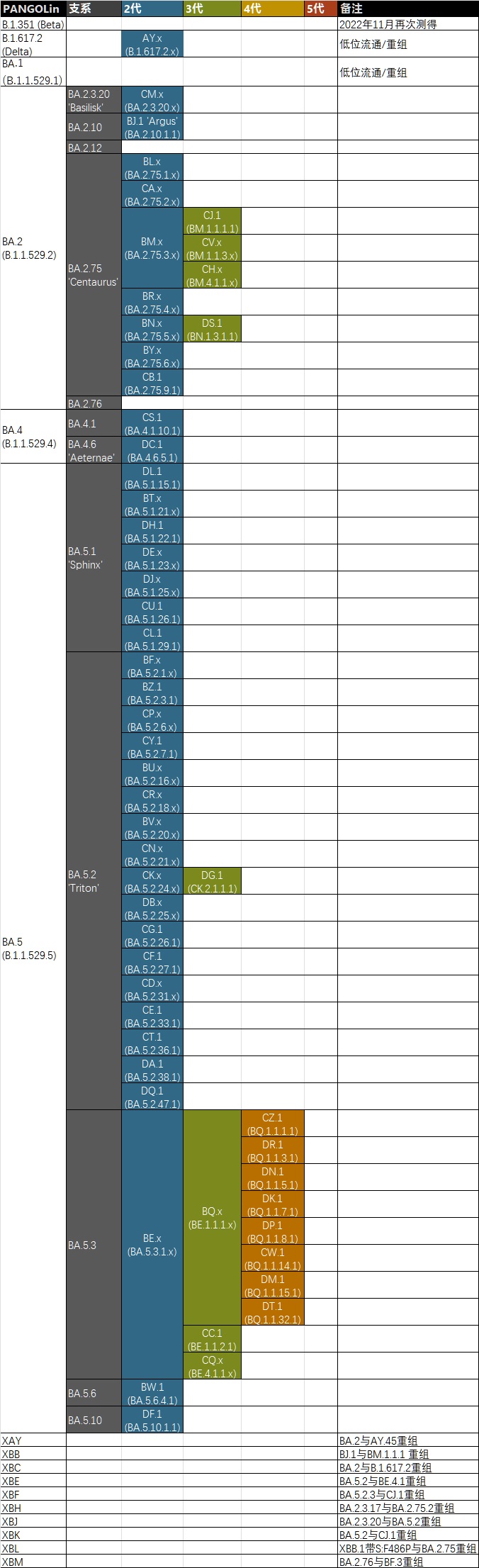
目前流行的變種和家族、代次關係
製圖的時候還沒算最新的DU.1,即BQ.1.1.2.1
除了大家已經熟知的BQ和XBB家族以外,還應該注意一下陪跑了小半年的CH家族。目前它在歐美國家中流行,已經傳入香港,需要注意的是其中有一個分支CH.1.1.4獲得了S:P681R,這是消失了小半年的著名的Delta合胞能力、致病性和傳播性三增突變(靠近弗林切割位的精氨酸R突變似乎都有類似的能力)。值得慶幸的是目前這個分支是在富裕地區(列支敦士登和德國巴登符騰堡邊上的奧地利福拉爾貝格)發現的,及早引起了重視,但問題是歐洲的其它測序拉跨地區是否已經產生了類似的突變?此外它的另一個分支CH.3同樣獲得了S:F486P,子代CH.3.1甚至還獲得了S:S494P又升了一階。
獲得了Nextstrain系統22G提名(但是沒有通過)的BN家族仍然在緩慢爬升中,目前佔據了小半個稀巴國並向鄰國溢出(甚至想要誣賴到中國遊客身上,活幹得也太糙了)。其中一個分支BN.1.4.1開始在開放閲讀框(ORF)上整花活,獲得了ORF3a:P240S、ORF1a:T4175I,這些都是加強合胞能力、複製能力和顯著增加致病力的突變,也是消失了有一段時間了,最早出現在Alpha和未能來得及獲得WHO提名就被BA.1拍死的高致病性毒株B.1.1.519上。
重組株方面,XBF和XBK都是有S:F486P的高階大佬;XBL更是第一個被命名的重組株的再重組,由帶S:F486P的尚未找到源頭的某個XBB.1分支和BA.2.75組成;此外還有尚未命名的XBB.1.5與BQ.1.14的重組,以及XBB.1.5和CH.1.1的重組,王中王了屬於是。
此外還有德爾塔克戎重組株XAY和XBC,它們都在迅速開枝散葉,前者主要出現在歐洲和非洲,後者已經衝出亞洲走向了世界。假如它們的祖輩一個致病性高一個傳的快,那麼“雜交”了兩者精華部分的它們會不會又高又快呢?