好快的染色體變異!島上小鼠千年分化為六種_風聞
返朴-返朴官方账号-关注返朴(ID:fanpu2019),阅读更多!05-09 09:21
演化從未終止,篩選無處不在。
撰文 | 玉寶

迅速變異:千年形成六個新物種
在葡萄牙的馬德拉島,生活着六種長相相似的小鼠,可稱之為馬德拉鼠。它們雖然看起來非常相像,但卻存在生殖隔離,也就是無法在自然條件下相互交配,或交配後無法產生可育的後代,這符合生物學上劃分新物種的標準,因此這六種小鼠屬於不同的物種。
科學家研究發現,這六種小鼠是在過去的一千年當中分化而成;在一千年前,它們有着共同的祖先。2000年的一篇Nature論文[1]提升了學界對馬德拉鼠的研究熱度,馬德拉島隨後就像太平洋上的加拉帕戈斯羣島一樣,成為研究生物演化的另一個聖地。眾所周知,1835年參加遠洋航行的達爾文就是在見到加拉帕戈斯島上的象龜、達爾文雀等特有物種後,逐步形成了生物演化理論或進化論。

生活在馬德拉島上的小鼠有什麼特別之處呢?科學家發現,它們的基因數目差別不大,但染色體的結構和數目會有所差異。這説明,馬德拉小鼠的演化走的不是基因變異的路線,而是通過我們較少聽説的染色體變異(chromosomal variation)來完成的。普通實驗室小鼠(Mus musculus)的染色體數目是20對40條,而馬德拉鼠的染色體數目介於22-40條之間。大部分突變的馬德拉鼠很難產生下一代,要麼在胚胎期死亡,要麼在出生後因各種缺陷而死亡。少數能夠繁育且能適應當地環境的,就會把變異保留下來,也就是我們現在看到的六種馬德拉鼠。
羅氏易位:也曾造就人類
藉助染色體顯帶技術,科學家在實驗室可以輕易發現染色體的各種數目和結構變異。造成馬德拉鼠染色體差異的主要原因是發生了羅賓遜易位(Robertsonian translocation),簡稱羅氏易位。
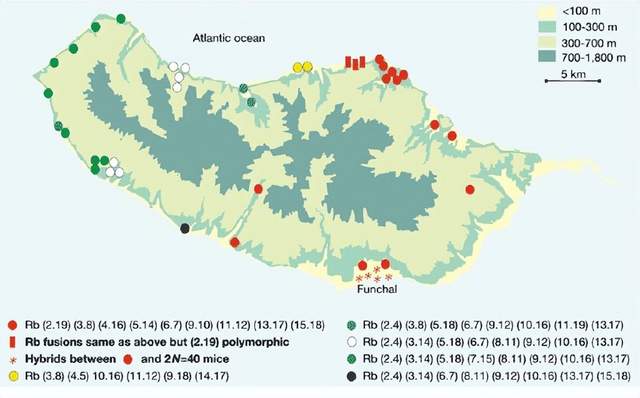
羅氏易位是指某兩條染色體的長臂融合成為一條染色體,並失去短臂。在細胞分裂的中期,有些染色體的着絲粒會位於染色體的一端,其短臂極短,這一類染色體稱為近端着絲粒染色體,羅氏易位往往發生在近端着絲粒類型的染色體之間。例如,人類的二號染色體,就是在數百萬年之前由兩條染色體發生了羅氏易位融合而成的[2]。據統計,羅氏易位在新生兒中的發生率是千分之一[3],大部分患兒會產生健康問題,如唐氏綜合徵和巴陶氏綜合徵。
馬德拉鼠的羅氏易位情況與人類似:在鼠羣中以一定概率發生,少部分不影響生殖和健康的易位被保留下來,進而逐步在種羣中擴散,甚至形成新的小鼠類羣。在這個740平方千米的島嶼上,目前確定有六種小鼠,它們的染色體核型明顯不一樣。可以想見,隨着採樣密度增加或時間推移,大概率還會發現/出現新的小鼠種系。
據推測,島上的小鼠祖先來自一千年前在此停靠的維京海盜船,另一個説法是隨15世紀葡萄牙殖民者的船而來。在現當代,隨着船舶或壓艙水而蔓延到世界各地的動物造成的物種入侵案例,也是數不勝數。
回顧中學生物知識,1個卵母細胞經過兩輪減數分裂,形成了1個卵細胞和3個極體。極體的命運是退化消失,只有卵細胞才會參與受精過程和個體發育。中學教科書寫着,物種染色體突變的特點是無偏好、隨機發生,後代比例符合孟德爾遺傳定律。但馬德拉鼠的染色體分離並不如此:在雌性小鼠減數分裂過程中,羅氏易位大概率會藉助卵細胞傳給下一代,而不是進入極體走向“絕路”。因為極體的命運是退化或死亡,並不參與受精過程及個體發育。
那麼,為什麼發生了羅氏易位的“大染色體”(兩個長臂融合,染色體體型較大)傾向於進入卵細胞而不是極體?
答案在着絲粒上。着絲粒是真核生物細胞中連接紡錘絲和染色體的結構。研究發現,羅氏易位後形成的染色體往往DNA含量更高,附着了更多的着絲粒蛋白,因此“大染色體”更容易分配到卵細胞中而不是極體中。
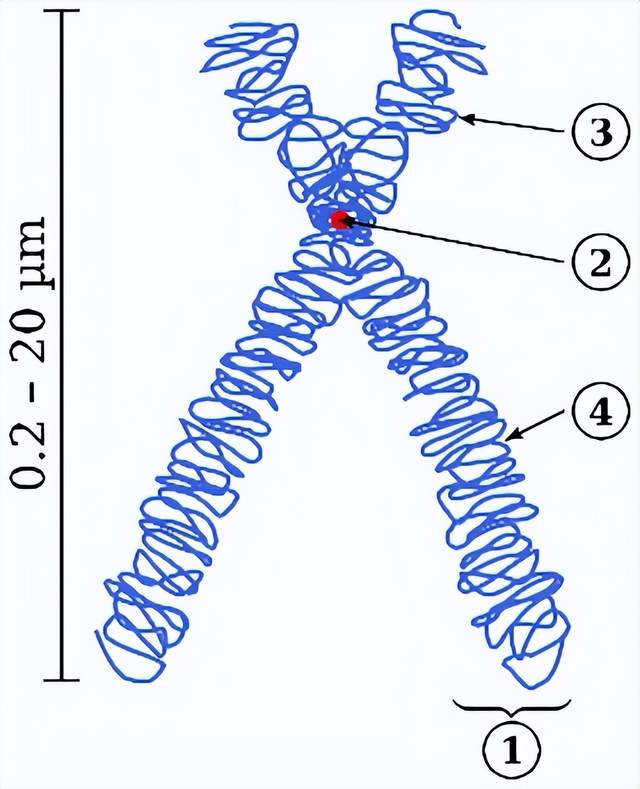
①染色體分節 ②中節,將染色體分為二臂。中節包含着絲粒(kinetochore),着絲粒是一種複合蛋白,將將染色體和紡錘絲微管相結合。③短臂 ④長臂丨來源:Wikipedia
自私染色體,善於“搭便車”
動物學家Michael J. D. White在50多年前曾提出,羅氏易位善於搭減數分裂的“便車”而傳下去。他認為,“少數染色體重排——例如羅氏易位——能夠在物種形成中發揮作用,在雌性的減數分裂過程中產生特定的優勢,進而通過這些機制形成生殖隔離”。馬德拉鼠染色體變異的機制證明了他的理論。
植物中是否也有類似的現象?
實際上,在馬德拉鼠之前,科學家就發現玉米和黃色猴面花等植物中具有大染色體的表型。

1942年,遺傳學家Marcus Rhoades發現[4],玉米的10號染色體有一個普遍發生的變異——Ab10,也是由羅氏易位形成的。具有Ab10突變染色體的玉米棒以黑粒玉米為主,間雜少量黃粒,而黑粒玉米棒出現的概率遠遠高出隨機發生的比例。對此,Rhoades提出的解釋是:
帶Ab10的染色體更容易進入到發育成卵細胞(而非極體)的位置,因此易位的後代個體數更多,其後代比例不符合孟德爾定律。現在我們知道, Ab10的確在減數分裂過程中更容易進入卵細胞,這和馬德拉鼠的羅氏易位是相似的。
Rhoades的發現是歷史上對“減數分裂驅動”現象的第一例報告。所謂減數分裂驅動,是指機體的染色體在減數分裂時因自私基因干擾, 後代比例偏離了孟德爾遺傳的期望比例。受“自私基因”(selfish gene)概念啓發,人們把玉米的10號染色體稱為“自私染色體”。直到2018年,才有一篇論文解釋清楚了Ab10的發生機制,揭示了參與這一過程的核心蛋白KINDR以及它的功能[5]。
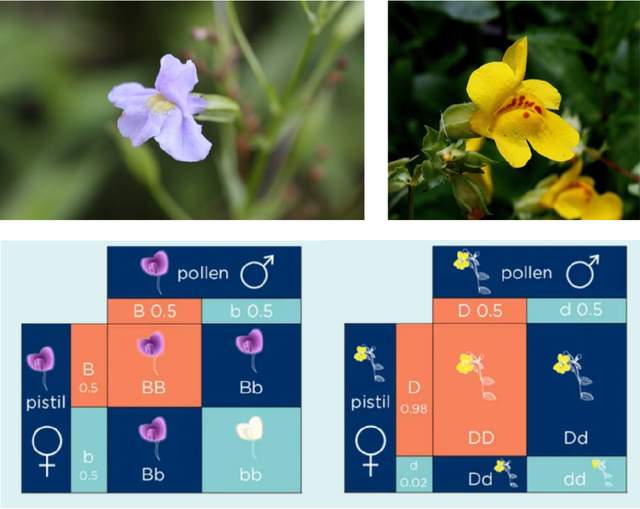
黃猴面花 (Mimulus guttatus) 的遺傳是減數分裂驅動(Meiotic drive)理論的另一個例證。2008年一篇論文表明[6],大着絲粒的黃色猴面花個體會產生更多後代,甚至顯性純合後代比例佔實驗羣體總數的50%左右,而按孟德爾遺傳定律這一比例應該是25%才對。參與這一機制的分子有CENP-A和CenH3等。
頻繁的變異:為自然選擇提供素材
通過上述案例,大家知道着絲粒在減數分裂中扮演的角色十分重要,沒有它,減數分裂、有絲分裂都無法進行。那麼,從常理出發,着絲粒的基因序列和蛋白在演化上應該非常保守(不輕易發生改變)才對,但事實是,真核生物的着絲粒蛋白、以及基因都不保守,變異非常快。
為什麼會發生這種悖論?2001年有科學家解釋説[7],是因為着絲粒參與了減數分裂驅動。根據這一解釋,着絲粒的基因序列——如玉米的Ab10——想“綁架”了行使染色體分離功能的“機器”,使這個“機器”不斷產生染色體變異,例如產生染色體非整數倍的卵細胞。變異的“危害”有大有小、也有中性突變,並非都是致死性的,所以這一過程就一直在快速演化,而沒有從羣體中消失。
在小鼠中,減數分裂驅動現象同樣存在。科學家發現,具有GTP酶活性的Ran和Cdc42的表達產物(蛋白質)參與了減數分裂驅動,影響到紡錘體的定位和極性建立[8]。通過把“大着絲粒”表型與“小着絲粒”表型的小鼠雜交,科學家發現,含“大着絲粒”鼠的染色體更容易進入卵細胞而不是極體。
演化從未終止,篩選無處不在。對於機體演化適應性,減數分裂驅動或“自私染色體”有什麼意義呢?
至少有一點是明確的:通過調控着絲粒與紡錘體微管相互作用,同源染色體會出現功能上的差異,為自然選擇提供源源不斷的素材。唯有如此,當自然環境或人工環境變動不居時,總有一部分攜帶某些變異的生物可以適應,逐漸造就更多的品系乃至物種。
要知道,數百萬年前的一次羅氏易位使得人的祖先與猿的祖先分離,類人猿才逐步演化成當今的人類。
參考文獻
[1] Janice BD et al, Rapid chromosomal evolution in island mice. Nature 2000, 403(6766): 158
[2] Paweł Stankiewicz. One pedigree we all may have come from –did Adam and Eve have the chromosome2 fusion? Molecular Cytogenetics 2016, 9:72
[3] Song JP et al. A family with Robertsonian translocation: a potential mechanism of speciation in humans. Mol Cytogenet. 2016, 9: 48.
[4] Rhoades M. Preferential segregation in maize. Genetics 1942, 27:395–407.
[5] R. Kelly Dawe et al. A Kinesin-14 Motor Activates Neocentromeres to Promote Meiotic Drive in Maize. Cell 2018 173(4), P839-850
[6] Finseth Fret al. Selfish chromosomal drive shapes recent centromeric histone evolution in monkeyflowers. PLoS Genet 2021, 17(4): e1009418.
[7] Harmit S Malik et al. Conflict begets complexity: the evolution of centromeres. Cur Opin in Gene & Dev 2002, 12: 711–718
[8] Benoit D et al. Ran GTPase promotes oocyte polarization by regulating ERM Ezrin/ Radixin/ Moesin inactivation. Cell Cycle. 2013 Jun 1;12(11):1672-8.
本文受科普中國·星空計劃項目扶持
出品:中國科協科普部
監製:中國科學技術出版社有限公司、北京中科星河文化傳媒有限公司

1. 進入『返樸』微信公眾號底部菜單“精品專欄“,可查閲不同主題系列科普文章。
2. 『返樸』提供按月檢索文章功能。關注公眾號,回覆四位數組成的年份+月份,如“1903”,可獲取2019年3月的文章索引,以此類推。
版權説明:歡迎個人轉發,任何形式的媒體或機構未經授權,不得轉載和摘編。轉載授權請在「返樸」微信公眾號內聯繫後台。